



SCI期刊那么多,一旦选错,费时费力不说,千辛万苦找到一个期刊投稿后,收到“论文内容与期刊不一致”的拒稿通知,对作者来说也是个不小的打击。更有一些水刊...那么我们该如何又快又精确地找到与论文内容最匹配的期刊呢?今天就给大家推荐一个SCI论文精准高效投稿利器:JournalGuide,只需1s,搜索相似论文,查看最匹配SCI期刊,实现精准投稿!网址:https://www.journalguide.com/优势:1、覆盖面广 ...
撰写学术论文已成为当今高材生们的必修课,除了大名鼎鼎的中国知网之外,你还知道哪些好用的论文网站呢?这次小编给大家推荐几个超 nice 的论文网站,方便大家在撰写论文时能更快地获得相关资料,用于借鉴参考。1. 师大云端下载站, 中文论文、专利检索下载链接:lunwenyizhan.com【师大云端】提供一站式论文、专利下载服务,面向全社会开放,国内网络都支持,校内校外都能高速下载。提供包括中国知网在内的八大文献数据库资源下载服务,数 ...

① 组蛋白修饰「阅读器」ZCWPW1 在雄性减数分裂中有决定性作用不孕不育在我国育龄夫妇中的发生率已达 15%,配子(精子或卵细胞)不能产生或其质量低下是引起不孕不育的重要原因,而精子和卵细胞必须经过减数分裂才能产生,一直以来,减数分裂的分子机制及存在的雌雄差异是研究的热点。来自香港大学深圳医院生殖医学研究中心,山东省立医院等处的研究人员发现组蛋白 H3K4me3 修饰「阅读器」ZCWPW1 对雌雄两性减数分裂具有不同效应,Zcwpw1 ...
① 北京基因组所 Autophagy 揭示去泛素化酶 USP33 调控线粒体自噬新机制PINK1 -Parkin 介导的线粒体自噬在线粒体质量控制过程中发挥着关键作用,其调控异常与人类神经退行性疾病发生相关。已有研究表明 Parkin 蛋白泛素化和去泛素化修饰参与线粒体自噬调控过程,但 Parkin 蛋白的去泛素化酶及其调控线粒体自噬的分子机制尚不清楚。中国科学院北京基因组研究所赵永良研究组首次证明去泛素化酶 USP33 定位于线粒体外膜,利用质谱分析 ...
① 华中科技大学最新研究:免疫机制参与心肌梗死后缺血性心力衰竭华中科技大学同济医学院附属协和医院心血管内科程翔教授研究团队发现了缺血性心力衰竭患者心脏局部 T 淋巴细胞免疫组库特征及免疫功能表型,在缺血性心力衰竭的免疫发病机制领域取得重要进展。这一研究团队发现在梗死后心力衰竭患者的心脏局部存在显著的 T 细胞应答;心脏局部的 T 细胞具有克隆扩增和组织特异性的表型,分泌促纤维化及促凋亡因子参与不良心室重构,且能再次进入外周循环;C ...
今天小编为你介绍常用的生信分析工具,这对你撰写研究方案和SCI文章写作很有帮助哦!1. solQTL (了解分析茄科、菊科的表型和基因型)http://solgenomics.net/qtl/ solQTL是番茄基因组测序计划的基因组信息资源门户网站数据库SGN (http://solgenomics.net) 的子模块。在solQTL上可看到全基因组QTL图谱的很多参数资料: 同时,还能清楚看到染色体每个位点: 目前的植物数据库Gramene ( ...

今天给大家安利一个好用的数据库:CircInteractome(Cirular RNA interactome)可以说功能很强大了,下面我们来看看它有哪些功能吧。一、搜索 circRNA 的名字、序列、在基因组上的位置及结合的蛋白首先是 circRNA 的搜索:我们可以根据 circRNA 或者基因名进行搜索,比如以基因 Cul2 为例,在 Gene Symbol 后面的对话框中输入 Cul2,然后单击 circRNA Search:在新页面中就出现了这个基因来源的各个 ci ...
circRNA的数据库还是很多的,以下几个可供参考:1.circBase收集包括以下6个物种的circRNA信息:人 (hg19)、小鼠(mm9) 、秀丽线虫(ce6)、黑腹果蝇 (dm3)、矛尾鱼(latCha1)、腔棘鱼 (latCha1)。该数据库是目前相对收录信息比较全和完整的,数据也能下载,非常实用。2. Circ2Traits是一个收集与人类疾病或性状潜在关联的circRNA数据库。3. circNet,利用464个RNA-seq测序 ...
① 傅嫈惠院士发现 ADRB1 基因能缩短人的睡眠时间!美国科学院院士傅嫈惠,和加州大学旧金山分校神经学家 Louis Ptáček 研究分析了几个家族,他们的睡眠时间只需要 6 个小时,而针对这个家族,研究人员发现了一个名为 ADRB1 的基因,在遗传连锁研究和全外显子组测序研究中,他们发现了这种新颖且非常罕见的突变。研究发现,这个基因编码ß1 -肾上腺素能受体,而这个蛋白的突变体版本稳定性较差,改变了受体的功能。这表明它可能在大脑中产生功能性后果。 ...
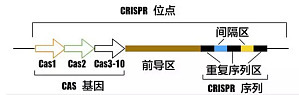
CRISPR 基因编辑技术自问世以来,就有着其他基因编辑技术无可比拟的优势,其应用在医学科研领域后,也不断的取得喜人的成果,进而迅速风靡于世界各地的实验室。那么 CRISPR 技术如此受到科研界的青睐,到底有什么过人之处呢?本文则揭开 CRISPR 神秘的面纱,让你快速掌握该基因编辑热门技术的精髓所在。CRISPR 原理及基本结构CRISPR/Cas 系统作为原核生物的免疫系统,可识别外源性遗传物质 DNA,并将其切断,沉默外源基因的表达。在 ...

大多做科研的童鞋们大概都会遇到一个头疼的问题:怎么找文献?如何保证找到的文献都是相关领域的经典文献?本文教你如何根据 H5 指数查找相关领域的高精尖经典文献。首先来了解一下什么是 H 指数、H5 和 H5 中位数?H 指数(H-index):于 2005 年由美国加州大学学圣迭哥分校物理学家 Hirsch 教授提出,用于评价个人学术影响。Hirsch 将该指数定义为:若某位科学家的 Np 篇论文中有 h 篇论文每一篇的引量都至少为 h 次,且其他(Np-h)篇论文中每篇的引量都
小鼠灌胃方法比较简单,需要关注的只有两点:一是要保持小鼠的头部和颈部成一直线,方便灌胃针头进入;二是动作要轻柔,从口角进入,防止损失食道。做的多了自然就熟练了。具体操作过程如下:1. 准备灌胃针头。一般可以从市场上面买到,实在没有的话,可以用 12 号的针头,剪去针尖,用砂纸将头端磨平,也可以用。但是买的灌胃针头的头端用锡或者适宜的方法处理了针头的锐口,自己用砂纸不可能将所有的锐口都磨掉,用这样的针头灌胃,损失小鼠食道的可能性比 ...
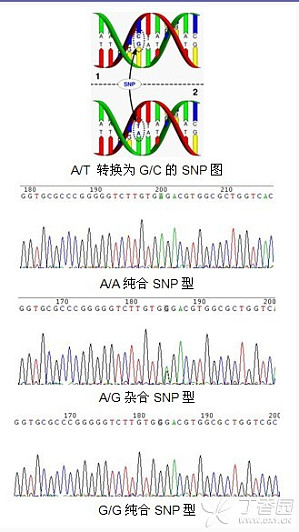
SNP,全称Single Nucleotide Polymorphisms,是指在基因组上单个核苷酸的变异,包括转换、颠换、缺失和插入,形成的遗传标记,其数量很多,多态性丰富。从理论上来看每一个SNP 位点都可以有4 种不同的变异形式,但实际上发生的只有两种,即转换和颠换,二者之比为2:1。SNP 在CG序列上出现最为频繁,而且多是C转换为T ,原因是CG中的胞嘧啶常被甲基化,而后自发地脱氨成为胸腺嘧啶。一般而言,SNP 是指变异频率大于1 %的 ...
① 中国科大 PNAS 揭示 NK 细胞终末成熟调控新机制天然杀伤细胞(Natural killer cells, NK cells)是免疫系统的重要组成部分,在机体抗病毒和抗肿瘤中发挥重要作用。IL- 15 是调控 NK 细胞发育成熟、存活增殖及效应功能的关键分子,目前对 IL- 15 信号通路的负相调控及其对 NK 细胞成熟与功能的影响研究尚少。中国科学技术大学生命科学与医学部等处的研究人员发现 IL- 17A 通过上调 SOCS3 抑制 IL- 15 信号以阻碍 NK 细胞发育成熟,一方面提示 IL- 17 对于 NK 细胞的过度活化 ...
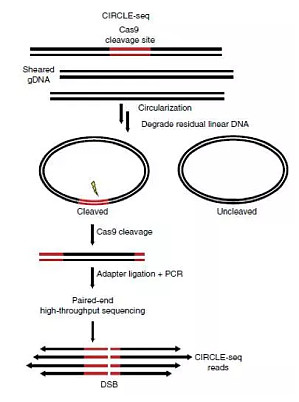
经典的 CRISPR/Cas9 系统具有较多优点,却存在很高的脱靶风险——不仅切割其靶向目标位点,在具有相似序列的位置也可能会发生切割 。下面给大家介绍一下 CRISPR 脱靶的检测方法。目前有很多可以设计 gRNA 并预测脱靶位点的网站,除了常用的 elevation 和 CRISPOR 外,还有 synergizing CRISPR 以及 deep CRISPR 等模型。但是如何从计算机的模拟预测中挑出一个完美 gRNA 呢?目前是 gRNA 切割后,测序检测其脱靶位点。但是还有预测工具 ...
一、选择荧光标记要匹配荧光直标抗体的一个主要优势在于它为多重检测提供了机会,比如现在一些先进的流式细胞仪能检测每个细胞中 20 多个离散参数。在设计多重实验时,应考虑每种荧光基团的独特性质,如最大吸收波长和最大发射波长,消光系数和斯托克斯位移等因素。对于免疫荧光分析方法,人们要同时检测组织或细胞样本中的多个抗原,常常会利用直标一抗进行免疫荧光共染色,这时一般尽量选择光谱重叠比较少的荧光基团进行组合。比如,我们会选择一个 ...
1、裂解物制备每道全细胞提取物的 20–30 µg 总蛋白,通常足够用来检测。如果靶标蛋白的基础水平,或蛋白质修饰程度低,可能需要通过化学刺激剂来诱导表达或修饰。你可能要调查备选细胞系或组织,其所含的目的蛋白含量更高。应将样品溶解于适当的缓冲液中,该缓冲液包含蛋白酶抑制剂和针对磷酸化目标的磷酸酶抑制剂。应务必对裂解物进行超声处理,以确保有效的染色质和膜结合靶标的蛋白质提取。请查看我们网站上的对照物表,获得推荐的阳性对照物和 ...
本方法免疫沉淀得到的天然蛋白可用于 western 免疫印迹或是激酶活性分析所需溶液和试剂注意:所有溶液用反渗透去离子水(Reverse Osmosis Deionized water, RODI)或相当纯度的水配制。1. 20 X 磷酸盐缓冲生理盐水(PBS): (#9808) 配制 1L 1 X PBS 时,取 50 ml 20 X PBS 用 950 ml RODI 水稀释到 1L,混匀。2. 10 X 细胞裂解液: (#9803) 配制 1 L 1 X 细胞裂解液时,取 100 ml 细胞裂解液加 950 ml RODI 水稀释到 1L,混匀。注意:使用前添加 1 mM PMSF。3. 蓝色上样缓冲液(S ...
免疫共沉淀(Co-immunoprecipitation)是以抗体和抗原之间的专一性作用为基础的用于研究蛋白质相互作用的经典方法,是确定两种蛋白质在完整细胞内生理性相互作用的有效方法。当蛋白质处于正常天然构象时, 某些抗体所识别的表位未能暴露出来, 这种情况下需要采取下列方法进行变性蛋白的免疫沉淀。所需溶液和试剂注意:所有溶液均需要用 Milli-Q 超纯水或是相当纯度的其他水配制。1. 变性细胞裂解液:50 mM Tris (pH 7.5), 70 mM β ...
① 禁食可以减少炎症,且不会降低免疫力!Cell 最新文章又一力证我们知道热量限制会改善炎症和自身免疫性疾病,但对于减少热量摄入,控制炎症的机制却知之甚少。西奈山研究人员利用人体和小鼠免疫细胞进行研究,结果发现间歇性禁食可减少血液循环中称为「单核细胞」的促炎细胞的释放。进一步的研究表明,在禁食期间,这些细胞会进入「睡眠模式」,而且比未禁食的人体内发现的单核细胞更少。这项新研究指出,短期和间歇性禁食可以减少慢性炎症,改善慢 ...

