




我还是在校生,正值看文献、做课题的时期。关于如何在短时间内获取文献全文、如何高效地从阅读的文献中获取我们急需的知识和内容,正是急需了解的,在此提出自己的一些拙见,不到之处,请大家海涵,指正。一、关于如何短时间内获得文献全文1、已知所求文献为哪一篇的情况下: 利用Google学术搜索引擎http://scholar.google.com Google真是个好东西,灵活运用,不敢说无所不能,但确实非常强大。Google学术搜索你所 ...
1. 选题选题是写好文献综述的首要条件。选题要从实际出发,具有明确的目的性,在理论或实践上有一定意义。选题来源包括:1)与自己实际工作或科研工作有关的、较为熟悉的问题;2)某护理问题的研究今年来发展较快,需要综合评价;3)从掌握的大量文献中选择反映本学科的新理论、新技术或新动向的题目。 文献综述的题目不宜过大,越具体越容易收集资料,从某一个侧面入手,容易深入。2. 搜集资料文献资料是撰写文献综述的物质基础,选定综述的题材后要大量 ...
文献综述是对某一方面的专题搜集大量情报资料后经综合分析而写成的一种学术论文, 它是科学文献的一种。文献综述是反映当前某一领域中某分支学科或重要专题的最新进展、学术见解和建议的它往往能反映出有关问题的新动态、新趋势、新水平、新原理和新技术等等。学写综述,至少有以下好处:①通过搜集文献资料过程,可进一步熟悉医学文献的查找方法和资料的积累方法;在查找的过程中同时也扩大了知识面;②查找文献资料、写文献综述是临床科研选题及进 ...
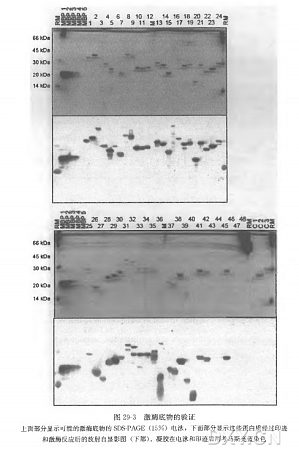
鉴定蛋白激酶底物是蛋白激酶研究的一个主要焦点,并为解析信号转导途径及其复杂的调节提供证据。我们在此介绍一种以蛋白质芯片技术为基础的离体实验方法,用特异蛋白激酶对固定在芯片上的蛋白质进行系统磷酸化筛选。这种高通量方法可以用来鉴定可能的激酶底物。这个方法从包含数百个纯化重组的组氨酸标记蛋白质的植物蛋白质芯片开始,要用可溶解的、有活性的激酶,在具有放射性的 ATP 存在的环境下孵育芯片。一次芯片实验只需要少量有活性的激酶。然后用磷屏成像仪或 X 射线胶片检测放射性信号。在活体内激酶与底物不能相互作用时,鉴定到的可能的蛋白激酶底物必须通过其他的体外或体内的方法进行确定。此筛选方法可以通用于鉴定多种蛋白激酶的可能底物的直接鉴定。
拟南芥和水稻基因组测序项目的完成极大地支持了蛋白质组学方法的应用,如蛋白质芯片技术。在本章中,我们介绍一种植物蛋白质芯片的构建方法及其在单克隆抗体或多克隆血清的特异性和交叉反应研究方面的应用。此方法从已鉴定的编码携带 His 标签的植物蛋白质的大肠杆菌(K cDNA 表达克隆开始,用高通量技术表达和纯化这些重组蛋白质后,用接触点样机构建蛋白芯片。用一种抗 RGS-His6 抗体来检测芯片上的这些重组蛋白质,为了分析特异性抗体,首先将芯片浸泡在各自的抗体溶液中孵育,接着在带荧光标记的第二抗体溶液中孵育。用芯片扫描仪检测信号。包含植物全蛋白质组的蛋白质芯片将是未来用以检测抗体的特异性和交叉反应的理想芯片格式。
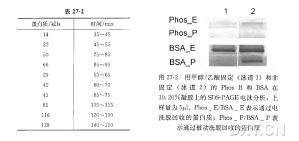
已有一种方法可用来从 SDS-PAGE 凝胶中提取完整蛋白质并对其进行 MALDI-TOF MS 分析,以确定其分子质量。这种方法包括一个电洗脱步骤以及在 MALDI-TOF 质谱分析之前的一个低温基质/ 分析物共结晶步骤。首先用电洗脱方法( ProteoPLUS )从聚丙烯酰胺凝胶中提取蛋白质。电洗脱之后,在相同的电洗脱管中,透析去除盐、SDS 和染料。真空离心浓缩蛋白质。以牛血清白蛋白 ( BSA )和磷酸酶 B( Phos B )做标准蛋白,结果发现,电洗脱比被动洗脱能更有效地从凝胶中提取完整蛋白质。优化后的蛋白质回收效率分别为:BSA 89% 和 Phos B 58%。此结果支持了回收效率与蛋白质大小负相关的假设。传统使用甲醇和乙酸“ 固定胶”的方法被认为会显著降低蛋白质回收效率,如果可能应避免这么做。下一步包括全蛋白与 MALDI 基质(反 3,5-二甲氧基-4-羟基肉桂酸,齐子酸,10 mg/ml) 低温共结晶的样品制备过程中也要进一步去除杂质。蛋白质和 MALDI 基质经混合、密封后贮存于 4℃ 过夜共结晶。然后除去上清液,再用相同的基质溶液溶解蛋白质-基质结晶。用低温共
环状 RNA(circular RNAs,circRNA)是一类具有闭合环状结构的 RNA 分子,早在 20 世纪八十年代即有研究报道,但由于其表达丰度低,文献报道较少,一直被认为是 RNA 转录剪切的罕见错误而被忽视。直到 2012 年开始有研究者开始大批量鉴定 circRNAs,从古生菌、线虫、小鼠和人类细胞鉴定出大量 circRNAs,揭示 circRNA 大量存在于真核转录组中,是细胞基因表达的一个普遍现象,并可能发挥重要的生物学作用。进一步研究表明 circR ...
高分辨率的蛋白质分离是蛋白质组学研究的一个重要条件,蛋白质的分离主要依赖于 2D IEF/SDS-PAGE,不幸的是,这种技术分离疏水性蛋白质的能力较差,而且不能用于研究自然状态的非变性蛋白质。蓝绿非变性聚丙烯酰胺凝胶电泳(PAGE)表现出一种可替代 2D IEF/SDS-PAGE 的蛋白质分离方法,这种方法基于用阴离子染料考马斯亮蓝将负电荷整合到蛋白质或蛋白复合物上,进行天然状态下的蛋白质分析。结合 SDS-PAGE 电泳,第一相蓝绿胶上被分离的蛋白复合物在第二相 SDS-PAGE 凝胶上被分离成其蛋白质亚基组分,在 2D 凝胶上形成竖条的点。2D 蓝绿胶 SDS-PAGE 是蛋白质组学研究中 2D IEF/SDS-PAGE 的理想补充技术。
原理聚丙烯酰胺凝胶是由丙烯酰胺(Acrylamide简称Acr)和交联剂甲叉双丙烯酰胺(N,Nˊ-MethyleneBisacrylamide,简称Bis),在催化剂核黄素或过硫酸铵和四甲基乙二胺的催化下,聚和而成的具有三维网状结构的胶。正常情况下蛋白的迁移率取决于它所带净电荷以及分子的大小和形状等因素。而在丙烯酰胺凝胶中加入阴离子去污剂十二烷基磺酸钠(SDS),则蛋白的迁移率主要取决于蛋白大小,与所带电荷与形 ...
题记科研之路,一入侯门深似海,从此悠哉是路人~实验之殇,雄关漫道真如铁,怎么老是撞到铁今天,老蔡就陪各位铁兄铁姐啃一块难啃的铁块-如何实现动物体内的基因转染?所谓基因转染,通俗一点说,主要指基因上下调(即 Gain of function OR loss of function)。基因工程小鼠-经典的动物体内基因干预模型最经典传统的动物体内基因干预,当属基因工程小鼠了。基因工程小鼠包括转基因小鼠,(TG mouse,transgenetic mouse ...
像其他真核细胞一样,在植物细胞中,糖基化是被研究最多的翻译后事件之一。根据寡糖部分和蛋白质骨架之间的连接方式,可将糖基化分为两种类型:N-糖基化和O-糖基化。在此介绍我们实验室常用的研究植物蛋白质糖基化的几种不同的实验方法。这些方法依赖于糖特异探针的印记检测、糖蛋白的特异去糖基化以及质谱分析。通过这些实验,不仅可以确定一个蛋白质是不是糖蛋白,还可以确定糖蛋白是在哪个位置以及如何被糖基化的。本章最后部分介绍植物细胞中糖蛋白类群的特殊纯化和鉴定方法,称为糖蛋白质组学。
可逆的蛋白质的磷酸化作用参与植物细胞生理学的所有活动。由于用于研究磷酸化蛋白质的蛋白质组学和质谱技术的应用,使得这项具有高度挑战性的任务—揭示和分析植物中动态蛋白磷酸化网络的工作最近才开始成为可能的。蛋白质胰蛋白酶的剪切,色谱层析分离富含磷酸化肽段,质谱裂解和磷酸化肽段的序列分析为建立鉴定大部分目前已知的植物体内磷酸化蛋白质的实验方法打下了基础。当用胰蛋白酶消化单独分离出来的组分或不同的植物细胞器的蛋白质时,这个方法是最有效的。固相金属螯合亲和层析(IMAC )对富集复杂的蛋白质消化物被甲基化后生成的磷酸肽很有用。以下分离出来的磷酸肽的串联质谱分析得到了磷酸化蛋白质的鉴定和生物体内磷酸化位点的定位。个体蛋白质中修饰位点的磷酸化程度的相对定量分析可借助稳定同位素标记技术或专用的液相色谱-质谱方法完成。

BioLegend MojoSort™ 磁珠分选实验分享:一、免疫磁珠分选简介免疫磁珠分选是一种从复杂的细胞混合物中分离出高纯度细胞的方法。基于细胞表面抗原能与连接有磁珠的特异性单抗相结合,在外加磁场中,通过抗体与磁珠相连的细胞被吸附而滞留在磁场中,不带有该种表面抗原的细胞由于不能与连接着磁珠的特异性单抗结合而没有磁性,不在磁场中停留,从而使细胞得以分离。有两种分选方式,阴性分选及阳性分选。BioLegend 公司的 Moj ...
PROTICdb 是一种主要为存储和分析通过 2D 聚丙烯酰胺电泳和质谱分析得到的植物蛋白质组数据的网络数据库。开发 PROTICdb 的目的是:①存储、跟踪、查询与蛋白质组学实验相关的信息,从组织取样到蛋白鉴定和定量分析;②将使用者的专业知识和其他来源的信息整合成一个用于支持数据分析的知识库(如用于等位基因变异或翻译后修饰产物的鉴定)。数据通过 Melanie,PDQuest,IM2d,ImageMaster(tm)2DPlatinum v5.0,Progenesis,Sequest,MS-Fit,Mascot 等软件处理之后可上传到 PROTICdb 关系型数据库中,也可以在网络上直接填写 ( 实验设计和方法)。2D 聚丙烯酰胺电泳注释图可以通过 GelBrowser 显示、查均和比对。定量数据可被方便地以列表形式导出,供任何第三方软件进行统计分析。PROTICdb 是基于 Oracle 或 PostgreSQLDataBase 管理系统,可从以下网址免费获取:http://cms.moulon.inra.fr/content/view/14/44/。

Preparation of Protein Extract1. Prepare extracts from cultured cells or tissues with our Trident Extraction Kits.The total number of cells per ml and the cell equivalent loaded per lane of gel should be optimized specifically for each protein and antibody.2. Determine the protein concen ...
用基质辅助激光解析飞行时间质谱(MALDI-TOF-MS)鉴定膜蛋白之前,需要将这些蛋白质分离出来用以质谱分析。用非变性去垢剂十二烷基-β-D-麦芽糖苷溶液增溶膜样品后,可用离子交换层析(IEC)分离膜蛋白质,并经十二烷基磺酸钠-聚丙烯酰胺凝胶电泳(SDS-PAGE),进一步分离蛋白质组分。根据膜蛋白质样品的复杂程度,可在 IEC/SDS-PAGE 之前,先使用凝胶过滤层析手段分离蛋白质。对最终获得的 SDS-PAGE 电泳胶板的染色,使我们能简单地建立起一个膜蛋白表达谱以及免疫抗原反应。此外,对疏水性膜镶嵌蛋白的胶内酶解也是很有必要的。通过上述这些分离步骤 ,最终可实现通过 MALDI-TOF-MS 对蛋白质的鉴定。本章介绍应用于植物、酵母质膜和植物液泡膜上的膜蛋白分离方法。
本章我们介绍运用 2D 液相色谱串联质谱大规模鉴定水稻叶片和根组织中的蛋白质的详细实验方法。该方法通过带有电喷雾发射喷器的纳升级流速双相层析柱( 强阳离子交换/反相)来实现,这种技术的缩写名为 Mudpit( 多维蛋白鉴定技术)。这里介绍的实验方法包括从水稻植株收集叶片和根组织材料,从收集的叶片和根组织材料中提取蛋白质,提取出来的蛋白质的消化,制备带有整合电喷雾发射喷头的双相毛细管层析柱,运行数据依据性二维液相色谱-串联质谱分离多肽,使用数据库搜索得到的串联质谱数据鉴定多肽和蛋白质。这个方法不仅适用于多种植物材料,而且可用于鉴定植物特定组织、器官、细胞器或亚细胞器的蛋白质。本章除了介绍详细的实验方法外,还列举了一个具有代表性实验的结果,即用多维蛋白鉴定技术鉴定超过 1000 个的从水稻叶片和根样品中提取的蛋白质。
(一)免疫(immunity)传统概念:指机体对病原体的感染有抵抗力。现代概念:机体免疫系统识别「自己」和「非己」,对自身成分产生天然免疫耐受,对非己异物产生排除的一种生理反应。免疫系统的功能:1. 免疫防御:识别、清除入侵的病原体及有害生物分子。2. 免疫监视:识别、清除体内突变细胞和病毒感染细胞。3. 免疫自稳:清除体内衰老、损伤或变易性细胞,对自身成分耐受,维持内外环境相对稳定。免疫的分类根据种系和个体免疫体系统的发育过程及免疫应 ...
串联质谱是根据肽段的裂解规律(fragmentation pattern,也叫做 MS/MS 扫描)对肽段进行鉴定的有效技术。在串联质谱中,每一个肽片段都可以获得各自的光谱 ,从而可以根据肽段的特性对蛋白质进行鉴定。这个在自动采集数据方面的重要优势给液相色谱和串联质谱建立了有效的联系,而纳升级色谱柱和电喷雾电离的使用可大大增加该方法的有效性。利用这种技术现在可以大量鉴定出飞摩尔(fentomole)水平的肽段。MS/MS 光谱的批量处理可以产生一系列肽段,这些肽段对鉴定纯蛋白或蛋白质混合物具有较高的可信度。
MALDI-TOF 肽质量指纹图谱(PMF)是鉴定蛋白质较为快捷、方便的方法,所研究的基因组已经完成了测序和注释,蛋白质可以通过二维凝胶电泳分离和检测。研究植物蛋白质组有两个困难:一是只有少量植物完成了基因组测序,二是主要污染物来自于非植物本身。本章介绍了经典的“自下而上”的方法(即从多肽到蛋白识别):凝胶切割,凝胶消化,肽回收和净化,MALDI-TOF 质谱,蛋白质数据库查询。

