




本文来自知乎(http://www.zhihu.com/question/20829666)1. 查找、阅读文献,用文献管理软件管理。一般一个完整的研究都是从查找和阅读文献开始的,通过阅读大量的文献,你才有关于做什么研究(主题)、怎么做研究(方法)和研究假设的想法。找文献,国内学校基本上是百链 (http://edu.blyun.com)+google scholar,国外学校就是学校图书馆+google scholar。然后文献管理软件推荐 Me ...
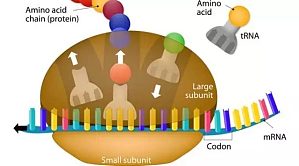
7 月 29 日,《自然》报道了分子生物学最新技术突破——基因工程师们开发出一种工具,通过控制细胞合成蛋白质的过程,让人们更好地理解蛋白质合成,探讨抗生素作用原理,并将细胞改造成合成化学物质的工厂。核糖体是细胞中的一种细胞器,除哺乳动物成熟的红细胞外,细胞中都有核糖体存在。一般而言,原核细胞只有一种核糖体,而真核细胞具有两种核糖体。细胞利用核糖体阅读转录自 DNA 的 mRNA 信息,利用氨基酸翻译成蛋白质。核糖体是细胞维持生存的基础,但是很 ...
免疫组化技术已经渗透到医学研究的各个领域,它在蛋白抗原的细胞准确定位上是其它蛋白检测方法无与伦比的。然而,要做好一张漂亮的染色切片,也不是十分容易的事。我做免疫组化方面的实验多年,积累了很多经典案例。下面我把这些案例列举出来,并附上我们发现的问题以及如何解决这些问题的解决方法,以供大家实验参考和实验交流。实验背景以前做免疫组化,我一直用中杉金桥的 SP 三步法染色试剂盒,效果不错。在奥运会期间我准备做同批实验分不同批 ...
经常看到别人论文中画出各种绚烂的插图,是用什么软件画出来的呢?分享一下我的绘画思路和常用软件。1. 绘画思路论文插图终归是内容导向,数据少/逻辑简单很难画出炫酷的图。专业图尽量用对应的专业软件画。大力出奇迹,插图最终效果跟投入的时间成本(学习软件+绘制+调试)成正比,只要耐心好学多尝试,哪怕 Office 系列也不会让你失望。绘图最重要的是专业性,其次关注「矢量+配色+字体+布局」。有时瓶颈在于思路而非工具,做科研的多浏览 Nature 和 ...
我最近两个月一直在做 ECL western,非常困难,都快做疯了。但是,功夫不负有心人,最终还是把困难的非特异条带和背景问题解决了。经过此番,我总结了几条心得与大家分享:第一条不要用平底小盒或杂交袋,用可以旋转的杂交管(可以用 50 ml 塑料离心管做成),我刚刚得到了比较结果,效果大不一样。第二条膜在 blocking 前先用 stripping buffer 处理一下(当然也在杂交管中进行,注意要通过旋转使处理均匀)。我偶然发现经过该处理后背景 ...
看了施一公院长在生科院毕业典礼上的演讲《大学的意义》,对一段话印象深刻:「我们清华人的奋斗目标从来不是、也不应该只是简单地找一份惬意的工作!我衷心希望你们每一个人在追求小我的同时,心里也要有一个大我——即便在困境,也要有一个承担起天下的雄心壮志。做事的时候要做到极致,不留下遗憾。生命就是体验,既然体验只有一次,何不做到极致?!」我想起了浙大老校长竺可桢的两个问题:诸位在校,有两个问题应该自己问问,第一,到浙大来做什么?第 ...
国际知名衰老期刊《Aging》近日发表了名为「 Bacopaside I 通过免疫调节的清除作用改善 APP/PS1 小鼠的认知损伤「的文章,该研究由第二军医大学药学院张卫东教授课题组完成 (Bacopaside I ameliorates cognitive impairment in APP/PS1 mice via immune‐mediated clearance of β‐amyloid )。目前 阿尔茨海默症(AD)是世界范围内最常见的神经退行性疾病之一。该疾病的发病机理极为 ...

做科研需要不少外文文献。一般来说 Elsevier,Springer ,知网,万方数据库等文献挺多,但是有时还会碰到查不到的文献,而这些文献确实需要,怎么办呢?1.Google 学术首先在 Google 学术搜索里进行搜索,里面一般会搜出来你要找的文献。在 Google 学术搜索里通常情况会出现「每组几个」等字样,然后进入后,分别点击,里面的其中一个就有可能会下到全文。当然这只是碰运气,不是万能的。同时,大家有没有发现,从 Google 学术搜索中,还 ...
自己做过不少实验,有些心得方法总结一下分享给大家。病毒滴度的测定常见方法有以下两种:稀释计数法滴度单位:TU/mL,指每毫升中含有的具有生物活性的病毒颗粒数。「TU」为「transducing units」的缩写,中文为「转导单位」,表示可以感染并进入到靶细胞中的病毒基因组数。第一天 细胞准备将生长状态良好的 293 T 细胞消化计数后稀释至 1×100 000/mL,加入 96 孔板,100 μL/孔,为每个病毒准备 10 个孔。放入 37℃,5% 二氧化碳培养箱中培养。第二天 ...
厌氧消化被认为是一种最重要、有效稳定的处理和回收污染的活性污泥,牲畜粪以及其他有机废水和固体废物的方法。有机物质可以通过厌氧消化过程被最大程度的转化成沼气,但是,剩余在沼渣沼液或发酵液中的矿物质和其他的中间产物比如挥发性脂肪酸却很难得到妥善处理。
看了标题点进来的,想必是想学得师姐一招半式,好在实验中大展拳脚。不会让大家失望的,今天就谈谈分子克隆的独家经验,可以回复目录查看以往师姐传授的经验。务必做好对照一定要做好对照,各种对照,只要你能想到。做的时候千万不要偷懒。比如构建质粒,我最后转化会至少分三个平板,一个涂实验组,一个涂连接过程不加酶的对照(用于估计酶的活性和转化的阳性率),一个涂不加质粒的感受态细胞(用于确定感受态本身没有被污染,或是抗生素没有失效)。 ...
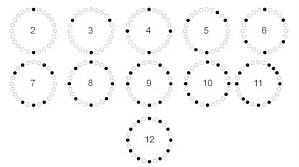
几乎每个做实验的朋友每天不知道要离心多少次,其中有一步配平,你真的会吗?现在考你一个问题,多半会答错。给你 n(n 取值为 1-24)个离心管,每个离心管内液体一样。这 n 个管都需要离心,且没有额外的配平管,有一个 24 孔离心机,请问 n 有多少种情况下不能离心?相信大家使用离心机时,都会遇到这样的问题:需要用 24 孔离心机离素数个管子。一般的做法是要再取一个离心管加水称重配平,这有些麻烦。偶数个自然,偶数个管肯定能配平,堆成放两边,所以偶数就不用考虑了。 ...

算算今年,总共有 11 个日本人获得诺贝尔物理奖。日本的物理科研这么强,对我们做生物科研有何借鉴呢?在我的研究生期间,辗转于北大,日本 KEK,和美国的 BNL。接触了三个不同地方的科研环境,相差还是非常的大。北大实验室给我打下扎实的基础,在日本 KEK 学到了努力和坚持是多么得重要,美国 BNL 给了我充分的自由和想象的空间。1. 日本科学家每天亲自做实验我在研二的时候去了日本 KEK 的国际直线对撞机部,参与超导加速器的预研。野口修一博士是我当时的导师 ...
细胞计数,看起来简单,其实暗藏玄机。今天就一起来学习下如何计数。1.原理计算细胞数目可用血球计数盘或是 Coultercounter 粒子计数器自动计数。血球计数盘一般有二个 chambers,每个 chamber 中细刻 9 个 1 mm² 大正方形,其中 4 个角落之正方形再细刻 16 个小格,深度均为 0.1 mm。当 chamber 上方盖上盖玻片后,每个大正方形之体积为 1 mm²×0.1 mm=0.0001 mL。使用时,计数每个大正方形内之细胞数目,乘以稀释倍数,再乘以 10000,即为每 mL 中之细 ...
做 MTT 实验容易走弯路。此前我们汇总过部分 MTT 实验相关细节,可点击阅读原文查看。今天继续和大家一起来谈谈MTT 实验步骤贴壁细胞相关实验操作1. 收集对数期细胞,调整细胞悬液浓度,每孔加入 100 uL 铺板使待测细胞调密度至 1 000-10 000 孔(边缘孔用无菌 PBS 填充)。2. 5% CO2,37℃ 孵育,至细胞单层铺满孔底(96 孔平底板),加入浓度梯度的药物。原则上,细胞贴壁后即可加药,或两小时,或半天时间,但我们常在前一天下午铺板,次日上午加药。一般 5-7 个梯度。每孔 100 uL ...
我做不出来克隆的时间长达半年,期间克隆实验方面许多问题都遇到过,现在总结主要的一些问题和大家交流,希望大家以后不要犯我一样的错,少走弯路。1. 感受态要验证感受态细菌本身有质粒污染,这个问题困扰了我三个月,一直到最近才找到原因,我这里犯了最大的错是没有做过感受态检测。也许大家奇怪为什么在三个月内的时间里没有怀疑到感受态,不用奇怪,因为感受态「表现的太正常了」,每次涂板都长菌斑,每板 20-30 左右,很正常。我一个同学用这样的 ...
跑电泳的时候,也许很多人会羡慕别人的电泳条带清晰好看,自己的电泳条带有时候弥散,有时候歪歪扭扭。熟能生巧,只要你掌握正确操作步骤,你也可以跑得得心应手。清洗玻璃板蘸点洗洁精轻轻擦洗。两面都擦洗过后用自来水冲,再用蒸馏水冲洗干净后立在筐里晾干。若不继续使用,需用无水乙醇擦拭后晾干再妥善收起来,玻璃板之间垫玻璃纸隔开。梳子应用水洗干净,临用前用无水乙醇擦拭晾干。灌胶与上样1. 玻璃板对齐后放入夹中卡紧。然后垂直卡在架子上 ...
据统计,约有 30% 细胞系被交叉污染或错误辨识,因使用了交叉污染或错误辨识的细胞会导致研究结论错误、结果不可重复、临床细胞治疗失败等。不仅浪费大量时间、精力和金钱,还可能造成不可挽回的灾难性后果。为此,很多细胞库现在都要对提交来的细胞系进行鉴定,并对细胞系之间的交叉污染进行监测。
我的课题涉及 Transwell 侵袭实验,因此在这里和大家分享 Transwell 侵袭实验。实验用品1.Transwell 小室我用的是 ECM554,用完后擦去基质胶,再用胰酶和 75% 酒精泡,可以把膜洗得很干净,用前用紫外里外都照 30 min。处理方法:用棉签轻轻擦去胶和反面细胞,清水冲洗,超声清洗,低档,30 min,清水 3×5 min,蒸馏水 3×5 min,室温凉干,用前紫外线小室正面 3 h,反面 6 h,微波,低火 10 min×2。因为我买的是铺好胶的,所以没买 Matrigel,二 ...
第一次写论文是不知从何处下手,第一次投递论文依然是不知所措。今天就手把手详解论文投稿步骤及潜规则。投稿程序1. 选择合适的 SCI 期刊,结合专业知识,根据年度影响因子表和他人经验来综合选择要投递的期刊,并进入该期刊查询系统查询近年来的文章走向。2. 下载:进入每个杂志的首页,打开 submit paper 一栏,点击 Introduction 查看或下载即可。3. 稿件及其相关材料准备:Manuscript.doc、Tables.doc、Figures. ...

