



俗话说,兵欲善其事,必先利其器。掌握一些好的软件,可以让你的科研工作达到事半功倍的效果。今天随我一起来看看分子生物学常用的一些软件,主要包括分子克隆、实验记录、文献管理。Primer Premier 5Premier 是由加拿大的 Premier 公司开发的专业用于 PCR 或测序引物以及杂交探针的设计和评估的软件。主要界面有序列编辑窗口(Genetank),引物设计窗口(Primer Design),酶切分析窗口(Restriction Sites ...

我和夫人多次成本申请到 2005 年的国家自然科学基金(以下简称国基),要感谢从丁香园学到的「独家秘笈」的成功应用。为了感谢丁香园,我们呕血力创,推出我们的秘诀,提供给正在奋斗的人们。一、审时度势所谓审时度势,关键在于要搞清楚基金究竟要赞助哪些人,一般来说,国家设立基金,目的就是赞助那些能够在科研上有突破的人。判断哪些人能够在科研上有突破,这个就依靠评委来判断了,要知道国基从来不会雪中送炭,只会锦上添花。你要申请,首先要想,假 ...

日前,《每日邮报》报道了一位得了肝癌的小妹妹。她的爸妈为了能让她一边治疗,一边上学,为她配备了一台未来机器人——「替身」(可怜天下父母心)。佩顿的机器人代替她重返校园。(图片来源:《华盛顿邮报》)佩顿通过出现在教室里的机器人和小伙伴们一起上课(图片来源:《华盛顿邮报》)佩顿通过出现在教室里的机器人和小伙伴们一起上课(图片来源:《华盛顿邮报》)此事一出,全网哗然。不过如此令人震惊的装备,竟然在丁香园里隐秘了 1 年,要不是因为这次的「替身」门 ...

医学史上满是奇怪和可怕的实验,有些实验虽然取得了成功,但过程也许可怕且不道德。乙肝疫苗的发现当时,斯塔顿岛的智障儿童学校中肝炎肆虐。为了研究该疾病,儿科医生 Krugman 将感染者的排泄物拌进奶昔,然后喂给学生。对的,你没看错,这货把带病毒的大便混进孩子的饮品中,故意感染其他孩子。这个实验从 1963 年持续到了 1966 年,最终他发现了甲肝和乙肝是两种不同的疾病,并且因此而研制出了乙肝疫苗。后来, Krugman 声称他的实验取得过孩子父 ...
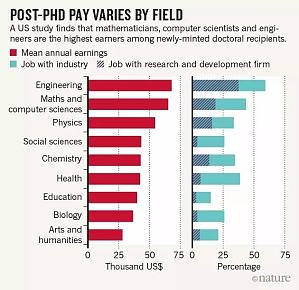
博士毕业后究竟该留在高校学术圈还是进入企业工作?哪里的收入更高?相信本文或许能为你今后选择提供一点参考。我们来看一项调查研究—— 关于 3 000 名美国博士生的经济情况。这项研究由纽约大学经济学家 Julia Lane 和俄亥俄州立大学经济学家 Bruce Weinberg 主持,项目由密歇根大学安娜堡分校发起,调查对象是来自美国中西部地区及周边大学的学生。其实研究的主要目的并非比较博士毕业生的收入情况,而是检验科学研究的社会经济生产力转化率。 ...

我是一间实验室,在这里已经第八年了。除了每天迎接清晨和夜晚之外,我能等的就只有一些穿着白大褂的人。虽然人不多,但是零零散散地进出也让我不觉得很孤单。实际上在这里有很多生命。我的孵箱里生活着亿万的细胞,只可惜他们很安静,连窃窃私语都没有。有时候一天天气很好,从早上就是一片云彩都没有,太阳就这么照射这里,投进屋子。这时候有人来,一边抱怨这屋里太热一边打开窗子通风。有时候天气不好,下了一夜的雨到早上都不停。我也不太开心遇 ...
看到有很多战友为引物的设计感到头疼。不过这也难怪,虽然原理我们都知道,但是设计的时候却未必能都考虑的很完全。在这里我向大家隆重推荐一个在线的引物设计软件,是斯坦福大学的,我认为是最好的在线引物设计软件,我用它设计接近 100 多对引物。它用起来也很简单,最重要的是效果非常好,考虑的因素也非常的多。软件操作页面:http://www.yeastgenome.org/cgi-bin/web-primer具体操作如下:首先输入你想要 p 的 ...
本文章主要提供给新手,希望对各位有帮助。1、引物是如何合成的?目前引物合成基本采用固相亚磷酰胺三酯法。DNA 合成仪有很多种,无论采用什么机器合成,合成的原理都相同,主要差别在于合成产率的高低,试剂消耗量的不同和单个循环用时的多少。亚磷酰胺三酯法合成 DNA 片段,具有高效、快速的偶联以及起始反应物比较稳定的特点。亚磷酰胺三酯法是将 DNA 固定在固相载体上完成 DNA 链的合成的,合成的方向是由待合成引物的 3' 端向 5' 端合成的,相邻的核苷酸通过3' ...
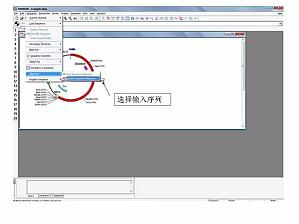
和大家介绍一下用 DNAMAN 软件制作适合论文发表用的 DNA 序列或蛋白质序列比对图。首先,选择输入序列 。选择序列文件所在位置。注意输入的序列是 DNA 或者是蛋白质序列,打勾相应的选项 ,然后一直点击下一步 。选项不需要修改,默认就行。由于是免费的版本,它会在这里加上了一段标记,需手工去除它。这是去除后的序列情况 。点击 Output ,出现下拉菜单。保持 EMF 格式的文件即可,然后用图片查看器打开可以放大缩小而不改变像素 。文章作者:丁香园论坛战友@gxu ...
前一阵子一直在做双酶切质粒重组,失败了很多次,不过很快改善了实验方法,用两周重组了 14 个质粒。现就自己的体会,结合丁香园战友的宝贵经验,谈一下质粒重组的一些个人经验。一、回收 PCR 产物在进行 PCR 扩增时候,给引物两端设计好酶切位点,一般说来,限制酶的选择非常重要,尽量选择粘端酶切和那些酶切效率高的限制酶,如 BamHI、HindIII,提前看好各公司的双切酶所用公用的 BUFFER,以及各酶在公用 BUFFER 里的效率。选好酶切位点后,在各 ...
和大家分享一下如何在新版 ensemb 查找启动子。首先,选择物种,输入基因名。比如: 搜索小鼠 Ptgs2 后,点击红圈处。点击红圈:点击 exons:5' upstream sequence 绿色部分为启动子所在位置,紫色部分是非编码区,黑色部分是编码区,蓝色区为内含子。但他所给的 5' upstream sequence 只有 50 bp,怎么办呢? 屏幕左边有个 configure this page,点击:更改数字 save and close,即可看到你想要的结果:Ensembl 除了可以用来克 ...
在互联网上,有很多软件可以解决分子生物实验人员在做实验中遇到的问题。本文提供了序列分析、实验设计阶段比较实用的软件。软件主要用于酶分析、引物设计、同源序列比较、质粒作图、结构域(motif)查找、RNA 二级结构预测、蛋白二级结构 分析、克隆策略图谱、三维结构显示等方面的内容。1. 综合性软件这类软件要求对本阶段上述的大部分功能均具备,但这类软件大多在专项分析上没有优势。推荐软件:Omiga 2.0优点:你所能想到的大部分对核酸蛋白 ...
这两年在美帝净做克隆实验了,专家不敢说,但是熟能生巧,确实积累了不少经验。我把经验写下来,希望大家少绕点弯路。1. 载体构建俗话说用欲善其事,必先利其器。我强烈建议大家在做构建之前先找好工具,这样起的效果事半功倍。这里说个笑话,我们系有个新 PHD 学生,是个印度女孩,很聪明很刻苦,她所在的实验室也很好,不过除了她之外包括老板在内都是生物物理背景的,以前一个生物 POSTDOC 在的时候还好,这个 POSTDOC 一走,整个实验室对分生就只有 ...
SCI 论文不好写,主要障碍在英语上,其实就像以前考四六级考研一样,撰写也是有规律可循,有模版可靠。Abstract越简单越好,字数控制在 150~250 words。具体怎么写呢?先简单描述一下课题的前沿背景,然后引出自己的实验(In this paper…. or Here, we…)主要陈述实验结果,可稍带提一下所用到的重要方法的名称,然后说明你的结果的意义 These data suggest….;最后总结拔高 In a word…. or In summary…..。举个栗子:Ac ...
俗话说,兵欲善其事,必先利其器。掌握一些好的软件,可以让你的科研工作达到事半功倍的效果。今天随我一起来看看分子生物学常用的一些软件,主要包括分子克隆、实验记录、文献管理。Primer Premier 5Premier 是由加拿大的 Premier 公司开发的专业用于 PCR 或测序引物以及杂交探针的设计和评估的软件。主要界面有序列编辑窗口(Genetank),引物设计窗口(Primer Design),酶切分析窗口(Restriction Sites ...
陆陆续续地回到实验室,该重操旧业啦。生物学霸今年会给大家呈上更多最新技术及解读,带你装逼带你飞。今天带给大家的是冷泉港二月期刊推出的最新实验技术,一共有 五个,学起来。CRISPR–Cas9 技术构建转基因鼠Editing the Mouse Genome Using the CRISPR–Cas9 System先用细菌举例给你解释一下啥叫基因编辑,比如我想让细菌给我产一个蛋白,但是他不听话,所以我只好人工地敲掉一些基因,然后再额外给他一些基因, ...
一般单次剂量的注射剂通过灭菌的方法即可达到无菌,而对于采用低温灭菌、滤过除菌或无菌操作法制备的注射剂或多剂量包装的注射剂,均应在处方中添加适宜的抑菌剂,以防止在制造、贮藏和使用过程中可能发生的微生物污染。抑菌剂具有抑制微生物细胞生长的能力,其过剂量也可能对人体有一定的安全性隐患,所以在注射剂处方中,抑菌剂的用量既要能抑制注射液中微生物的生长,又要求对人体无毒、无害。而对于加有抑菌剂的注射剂,仍要用适宜方法除菌, ...
著名俄国诗人 Paschkin 说过「灵感是在人们不断的工作中产生的」。因此,在你的科研活动之中,一定会产生许多的灵感,有些灵感信手拈来,有些灵感苦思而得,有些灵感受人启发。当你们抓住这些灵感并将它们付诸于实践之后,便有了许多精彩的发明和创新。一个小小的发明和创新为实验的顺利进行铺平了道路,我们收集了这些小发明,希望对你们有用。1. 病理实验dingle007:病理实验常需要专业人员看片,对炎症细胞浸润我做了个设想:即在同一组 ...
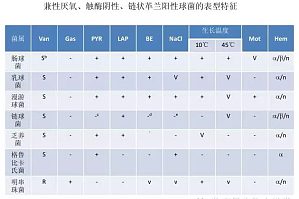
概述肠球菌属(Enterococcus)细菌域,厚壁门,芽孢杆菌纲,乳杆菌目,肠球菌科;在环境中广泛存在(土壤、水、植物、动物);人类胃肠道和女性的生殖道;不同种类的肠球菌的流行取决于宿主的不同,也受年龄、饮食和其他生理条件有关的因素影响。流行病学肠球菌是内源性和外源性医院感染的第三大病原菌(检出率仅次于金黄色葡萄球菌和大肠埃希氏菌)肠球菌所致院内感染绝大多数为尿路感染(有资料统计,在引起尿路感染的致病菌中,肠球菌感染 ...
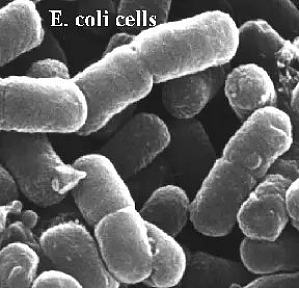
大家对大肠杆菌应该很熟悉,它是分子生物学里最常见的工程菌。基因克隆、蛋白表达,这些就是在大肠杆菌中实现的,这和大肠杆菌的结构密切相关。大肠杆菌 K12大肠杆菌是一种革兰氏阴性菌,棒状,约 2.5 微米长。有鞭毛,它的基因组由 4 639 221 个碱基对组成,至少编码 4 000 个基因。扫描电镜下的 E.coliK12 菌株最早是 1921 年从疟疾患者的粪便中分离得到,一直作为纯种保藏在实验室里,已有 75 年。大肠杆菌 K12 容易生长,并且关于它的代谢研究很多,因此它经常被用作 ...

