哺乳动物转录调控研究现状与展望
互联网
- 相关专题
不同细胞具有不同的表型,这是由于在这些细胞中基因的差异表达所导致的。因此,一定存在一些转录调控机制,去控制管理不同细胞各自所必需的转录方式。基因调控 这一领域的研究正是迅速地揭示转录的机制。不过,我们也必须要同事意识到转录调控的无限的复杂性。
正常的细胞进行正常的转录。但由于转录是基因表达 最基础的一级,所以当该转录调控出现异常时则很可能会出现不同的疾病。也正因如此,哺乳动物转录调控研究应用广泛于多种与人相关的疾病研究。近年来,其研究的方向主要集中于细胞通路的研究,并应用于发现疾病的发病或维持机制,从而最终选取适当的检测或治疗方法。
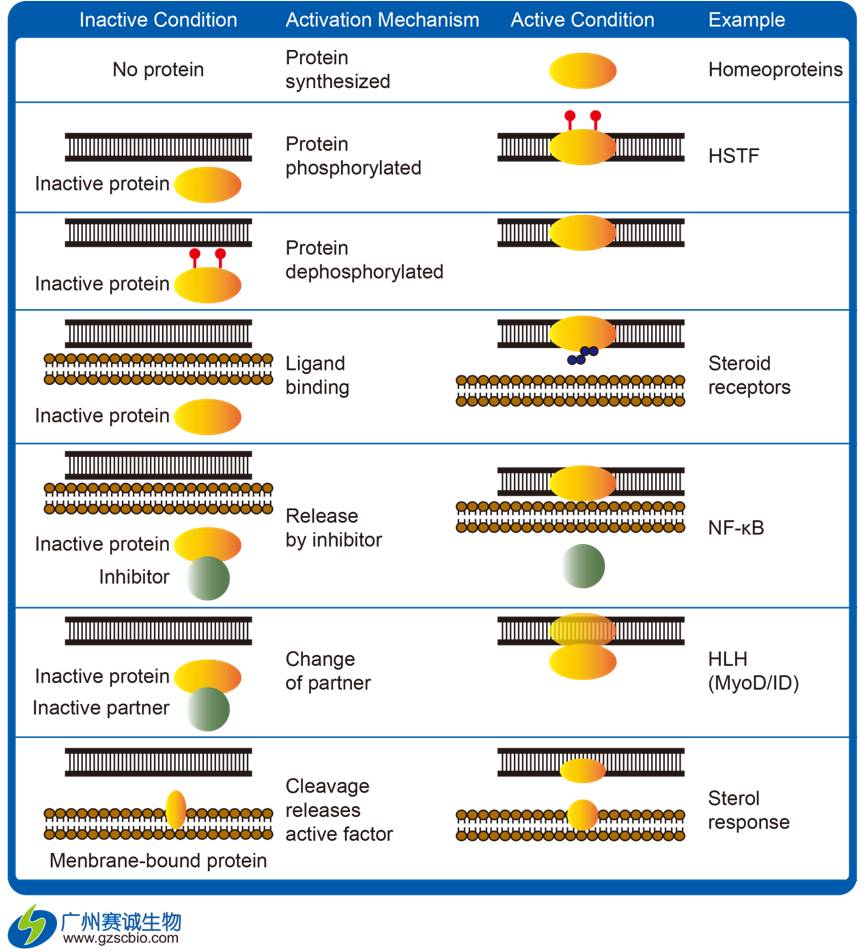
哺乳动物转录调控主要是在于DNA通过多种方法,使结构发生改变但不改变DNA本身的序列,最终影响其转录效率。这些方法主要包括DNA甲基化、组蛋白修饰、非编码rna 分子作用以及多种蛋白质结合修饰。其中,以最后一点尤为重要。
接下来,我们就哺乳动物转录调控(DNA与蛋白质互作 )这一专题,其研究现状及展望的论述。
一、哺乳动物转录调控研究的进展
1 转录起始的控制
1.1 rna聚合酶
转录其实是基因转录 的关键控制点,而关键的作用酶正是RNA聚合酶。RNA聚合酶是一类大分子蛋白质,是一个由8-14个亚基组成的复合物,其分子质量在500kDa以上。在哺乳动物的细胞中,转录通过三种RNA聚合酶中的一种得以实现,而每一种RNA聚合酶都具有不同的特征。
(1)RNA聚合酶 I转录rRNA,位于核仁,主要负责50~70%的RNA聚合酶活性;
(2)RNA聚合酶II合成核外RNA,位于核浆,主要负责20~40%的RNA聚合酶活性;
(3)RNA聚合酶III合成tRNA、5S RNA以及小核RNA,位于核浆,主要负责10%的RNA聚合酶活性。
1.2 基本转录因子(GTFs,General transcription factors)的装配
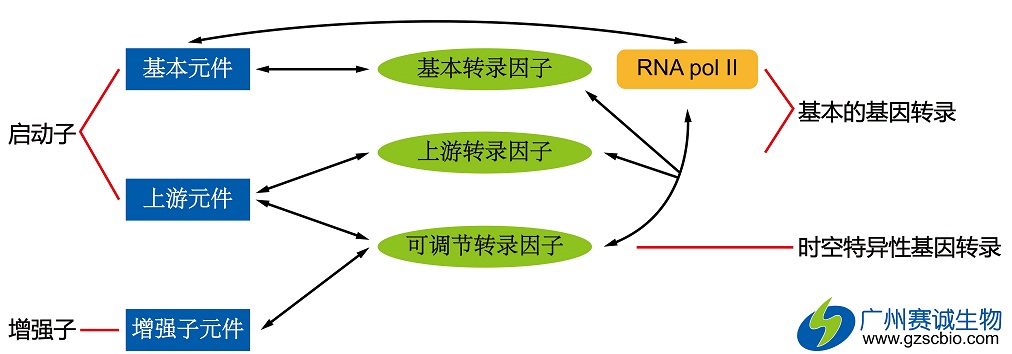
RNA聚合酶II主要负责哺乳动物的基因差异表达,这完全依赖于转录因子控制转录起始。RNA聚合酶II与辅助的转录因子的复合物就是我们常说的基础转录装置或GTFs。这些独立的部件GTFs聚集在近端启动子区域,提供了一个招集RNA聚合酶II的平台。随后,RNA聚合酶II就可以在一个精确的转录起始位点起始转录。
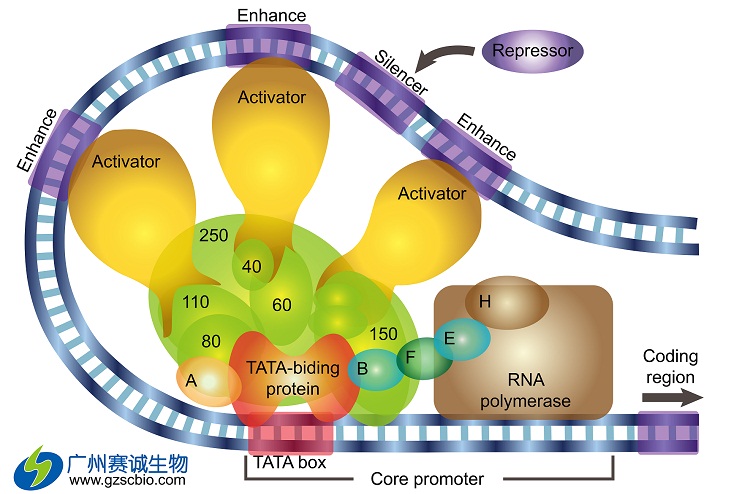
哺乳动物的启动子序列包含一段核心的保守的启动子。它可以产生足够的转录起始信号。在人类中,这段序列位于转录起始位点-45~-20bp的位置。而这段序列出现最多的情况是位于转录起始位点上游约25bp处的TATA box。TATA box被GC丰富的序列所围绕,并可以被TATA-结合蛋白(TBP)所识别。TBP属于GTF的一种,但也是唯一一种可以序列特异性结合DNA的GTF。TBP使得GTF的装配在TATA box上开始,并随后招集其他相关蛋白进行装配。在装配的过程中,RNA聚合酶II也会结合其中而开始转录,也可能会受增强子或沉默子等元件的影响,结合上其他蛋白从而调控整个转录过程。

哺乳动物转录起始GTFs的装配
1.3 非结构性GTFs的功能
在基础转录装置中,大部分装配的因子只被证实具有结构性的角色。但是,其中仍有部分因子已被证实具有酶的功能,作用于转录的起始,如TFIIE。
值得注意的是,对于不同的启动子,GTFs的装配以及转录的起始的RNA聚合酶是不同的。而且,很多的基因现在已经被证实具有无TATA的启动子。这些基因看来是依赖于上游的TFs起始转录的,如Sp1、ETS模体、富含嘧啶的起始模体。
2 转录调控起始的调节
对于任何一段候选基因片段来说,都需要一段核心启动子序列的存在以及基础转录装置的装配。但是,转录的水平实现与这样的一段启动子是最低限度的要求,而调控转录速率则需要其他上游因子。早期的一些构想暗示了,转录只有当候选基因被激活“打开”(TFs与启动子相互作用后的结果)之后才会发生。而现在,我们已经基本清楚了解到,除非转录速率被增强子增强或被沉默子“关闭”,否则基因会在其核心启动子作用下保持最低的速率持续表达。
3 增强子及其调控机制
很多增强子以及与它们相关的蛋白已被证实。而且我们现在已经清楚地了解到,一小部分增强子在大部分启动子中都很常见。如CCAAT box、GC box。这些元件对增加转录速率是十分重要的。但是,很多其他的模体也同样被证实对外来的刺激有反应,比且可以改变基因表达的速率,如“反应元件”。这些模体通常定位于较GC box和CCAAT box更远的上游端,但他们以同样的方式发挥着对转录速率的影响作用。
一开始,增强子都被认为是通过与RNA聚合酶II进行物理的相互作用而发挥其功能的。但是现在,人们已经相信它们的影响是直接的,这涉及到中介因子(将信号从激活因子传到转录蛋白)。通过一系列实验(如突变CTD,使得递质不能再与之结合,进而导致激活的蛋白的功能丧失),已经证实了这些递质作为中介因子而存在。
在一些早期的启动子结构及其功能的总结中,上游因子和反应元件局限于一些明确定义的基因区域,并按照明确定义的模式定向表达基因。然而,最近的一些研究表明,沉默子和增强子可定位于转录起始位点的5’或者3’端、内含子、外显子等,甚至是转录的RNA本身。直观地说,大部分的上游元件必须依赖于核小体的缩聚、DNA茎环结构以及蛋白质的柔性,而使得它们能与中介因子或者是顺式元件相互作用,进而最终改变转录起始及基因表达的频率。
二、哺乳动物转录调控研究的展望
从启动子结构、功能的特性及共性的相关知识中,我们可以知道在体内基因的表达是可被操作并最终受调控的。这提供给我们一个强大的工具的基础。
但到目前为止,仍存在许多仍未解决的问题。例如,在细胞有丝分裂的时候转录调控的明确机制如何?次生代谢的染色质结构及表观遗传代码是如何影响转录调控的?在一些特别的细胞(如ESC)中转录调控的共激活机制是怎么样的?
但先忽略这些未知因素,对于了解这些元件,更进一步的研究应更趋向于提供有价值的信息。通过结合实际的研究,我们在癌症及其他人类疾病的治疗中可更好地利用基于基因的疗法,也可以更好更全面地了解生物化学及生物分子学。
标签: 广州赛诚生物 哺乳动物转录调控
作者:Snail 点击: 次