




文章题目:Primase-based whole genome amplification文章出处:Nucleic Acids Research 2008 36(13):e79; doi:10.1093/nar/gkn377推荐理由:该文提出了一种不用加引物就能将基因组大量复制,你相信吗?请欣赏一下这篇精美的文章吧!设计的非常巧妙!摘要:In vitro DNA amplification methods, such as polymerase chain reaction (PCR), r ...
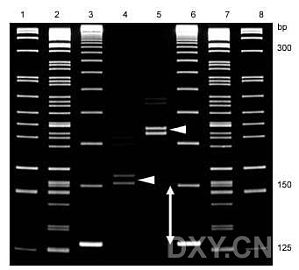
与大家分享一种高分辨率聚丙烯酰胺凝胶的制备方法——Enhanced 聚丙烯酰胺凝胶(EP凝胶)EP凝胶使用短胶板制备,一般10cm即可,制备后分辨率能达到1bp,并且核酸电泳条带比老方法制备的聚丙烯酰胺凝胶电泳条带更分散,更清晰。9%-12%的聚丙烯酰胺凝胶能达到1bp分辨率,6%-8%的凝胶能达到4bp的分辨率。Gel Conc.DNA Range bpAcrylamide-Bis, 40%, (29 : 1)Deionized WaterTAE Buff ...
【2005年3月】2005-03-14: Oxford BioMedica用RNAi慢病毒治疗家族ALS动物模型http://www.oxfordbiomedica.co.uk/news/2005-ob-09.htmhttp://www.nature.com/nm/journal/vaop/ncurrent/abs/nm1205.htmlOxford BioMedica (LSE:OXB.L), the leading gene therapy company, an ...
版主:最近小弟买了两套最新的《细胞培养技术》以及《分子生物学实验技术》的光碟,大约有1.1G,有需要的兄弟没有,如果有的话怎么样才能与大家共享?今天,我先把《分子生物学实验技术》光碟所包含的书名列在下面,请大家过目!1、《Molecular Cloning :A laboratory manual 》2、《PCR Cloning Protocols》Second Edition ,433页3、《PCR Mutation Detection Protocols》223 ...
(求助)原位杂交原位杂交?原位杂交(求助)原位杂交概念 谢谢:原位杂交流程原位杂交【求助】原位杂交 原位杂交收费?Re:求助!原位杂交(求助)原位杂交试剂盒【求助】原位杂交求助:HSP70原位杂交Re:(求助)原位杂交原位杂交求助请教原位杂交Re:原位杂交问题急请教原位杂交原位杂交掉片!Re:原位杂交?[求助]原位杂交试剂盒
忙了半天,整理完了,整理的不大好,多见谅1。基因测序的发展历史http://www.newlife.org.cn/xueshu/yantaohui/39-di3daijiyincexujishu.htm2。基因测序的几种技术非同位素银染测序系统操作技术(内容详细)http://www.biox.cn/content/labtec/20040824465.htmT7 DNA聚合酶测序技术(内容详细)http://www.biox.cn/ ...
标题:【请下载】Lehninger Principles of Biochemistry (35 Mb, 4th edition, 2005)内容:Lehninger Principles of Biochemistry格式:PDF大小:35 MB语种:英文简介:http://bcs.whfreeman.com/lehninger/default.asp---------------------------------------------------------------------------------------------------------Con ...
按一下步骤操作,连续两次均抽不出任何东西,请高手指点!抽提用作转染的重组质粒DNA(用5ml菌液)所用试剂盒为Promega公司的Wizard○R Purefection Plasmid DNA Purification System,能够去除对细胞有害的细菌内毒素。具体操作步骤如下:(1) 将5ml 培养过夜的阳性菌在室温(24℃)下10000×g离心10 min。弃上清,将离心管倒置至吸水纸上去除剩余水分。(2)加入250µl 重悬液,振荡至完全重悬 ...
1)1 M Tris-HCl (pH7.4, 7.6, 8.0)组份浓度:1 M Tris-HCl配制量:1 L配制方法:1. 称量121.1 g Tris置于1 L烧杯中。2. 加入约800 ml的去离子水,充分搅拌溶解。3. 按下表量加入浓盐酸调节所需要的pH值。PH值浓HCl7.4约70ml7.6约60ml8.0约42ml4. 将溶液定容至1 L。5. 高温高压灭菌后,室温保存。注意:应使溶液冷却至室温后再调定pH值,因为Tris溶液的pH值随温度的变化差异很大,温度每升高1℃,溶液的pH值大 ...
在凝胶成像系统电脑上发现的东西。园里不没有。呵呵。帖一个分享一下。园里相关的方法还有:酵母总RNA提取 mRNA的分离技巧小麦总RNA的提取验证有效的多酚类植物(苹果,棉花)提取RNA protocolRNA提取,欢迎对其细节进行交流土壤DNA、RNA的提取 RNA 操作注意事项与实验技巧操作注意事项: 由于RNA酶的广泛存在与难以灭活的顽固特性。使得RNA的提取纯化和后续工作变得非常困难。为了保证RNA的研究工作成功,请仔细阅读 ...
国家人类基因组北方研究中心对外开展重组腺病毒构建包装服务通过与美国Vector Biolab公司合作,国家人类基因组北方研究中心(北京诺赛基因组研究中心有限公司)利用优越的实验条件和训练有素的科研人员,成功构建的一个集克隆、包装、扩增、纯化为一体的重组腺病毒构建技术平台。从2005年7月成立腺病毒小组并开展实验至今,为课题组内部完成包装200多个腺病毒,为Vector Biolab公司成功包装近300个腺病毒,并且 ...
全基因表达谱差异研究有这些方法,EST测序(成本巨大)、DDPCR(特异性差,基本已经不用)、基因芯片(技术上比较复杂,数据处理工作量大)、SAGE技术(相对较科学,实验数据较可靠、花费较大)等。个人推荐:(本人原创,请斑竹加一分表示鼓励)cDNA-AFLP是从AFLP (amplified fragment length polymorphism) 衍生而来的RNA指纹识别技术,现已发展成较为成熟的转录组研究技术(Donson, et al., 2002) ...
核酸版有赖于yong主任、liven主任和管理员的倾心建设,以及后期加入的斑竹和各位战友的热情参与,凝聚了不少优秀的资源。为对这些优秀资源进行整理,以便于dxyer更好地利用本版资源,特向广大战友征集“旧帖整理”。有兴趣的朋友可以选择一个核酸基因技术方面的常规角度进行整理。我们将根据劳动量、整理视角的合理性、整理内容的安排的科学性等方面进行评价,赠予2分~5分不等的积分。好机会不容错过!核酸基因技术版期待因为您的 ...
看到这篇文章,非常的详细,和大家分享一下。由于内容比较多,编辑不是很方便,也上传了附件供大家下载。如果觉得有帮助记得投票或者顶贴哦!基因芯片 1什么是基因芯片 生物芯片,简单地说就是在一块指甲大小(1cm3)的有多聚赖氨酸包被的硅片上或其它固相支持物(如玻璃片、硅片、聚丙烯膜、硝酸纤维素膜、尼龙膜等,但需经特殊处理。作原位合成的支持物在聚合反应前要先使其表面衍生出羟基或氨基(视所要固定的分子为核酸或寡肽而定)并与保护基建立 ...
原位杂交的教程及问题分析,可以帮助分析杂交试验失败的原因.目录第一节 原位核酸分子杂交技术的发展?第二节 原位杂交技术的基本方法?(一)固定?(二)玻片和组织切片的处理?(三)杂交?(四)杂交后处理?(五)显示?(六)对照实验和结果的判断?第三节 原位DNA和DNA分子杂交方法?(一)地高辛(Dig)标记DNA探针在石蜡切片检测病毒DNA的方法?(二)生物素标记HPV-DNA探针在石蜡切片上检测HPV-DNA的方法?第四节 RNA原位核酸杂交方 ...
完整RNA的提取和纯化,是进行RNA方面的研究工作,如Nothern杂交、mRNA分离、RT-PCR、定量PCR、cDNA合成及体外翻译等的前提。所有RNA的提取过程中都有五个关键点,即1):样品细胞或组织的有效破碎;2),有效地使核蛋白复合体变性;3),对内源RNA酶的有效抑制;4)有效地将RNA从DNA和蛋白混合物中分离;5),对于多糖含量高的样品还牵涉到多糖杂质的有效除去。但其中最关键的是抑制RNA酶活性。RNA的 ...
到医院实习之前,我有幸跟随我的恩师,利用课余时间做了两年多的基础科研。从不懂规矩到渐入佳境,期间学习了很多东西,在此感谢我的恩师!我们是生物化学教研室,这两年的时间里我做了很多生化方面的实验。像细菌培养、细胞培养、PCR、western-blot、MTT测细胞增殖能力,这些实验都有商品化的试剂和推荐的操作方法,做起来,难度并不大。唯独让我感到苦恼的就是ShRNA表达质粒的构建这部分,它耗费了我们很多的精力,虽然最终 ...
现在许多从事分子生物学研究的人都非常恐惧EB,希望本帖对大家有帮助。EB只不过是被证明具有“潜在”的致癌性,并不表示一定就有致癌性!而之所以说是“潜在”的致癌性,仅仅是因为其致癌性是通过细菌诱变实验(Ames 测试)证明的结论。然而,用细菌进行检验的结果并不一定就能直接证明对人体或其它种类的生物具有相同的效果,这也就是为什么以前的《分子克隆》也只说EB有可能具有致癌性,而不是一定就是致癌物质的原因。Ames测试也只是对 ...
第一章 质粒DNA的分离、纯化和鉴定第一节 概 述 把一个有用的目的DNA片段通过重组DNA技术,送进受体细胞中去进行繁殖和表达的工具叫载体(Vector)。细菌质粒是重组DNA技术中常用的载体。 质粒(Plasmid)是一种染色体外的稳定遗传因子,大小从1-200kb不等,为双链、闭环的DNA分子,并以超螺旋状态存在于宿主细胞中。质粒主要发现于细菌、放线菌和真菌细胞中,它具有自主复制和转录能力,能在子代细胞中保持恒定的拷贝数,并表达 ...
最近做了四次电转化,两次成功转化率大于10的9次方,效果非常好 ;另从两次失败转化率吸取了教训;现将试验方案及操作与大家共享:1.细菌电转化感受态细胞的制备 (方案由李芃同学提供,lzf继承补充)1)由-70度冰箱中保存得菌种划单菌落,过夜培养16-20hr。晚上将将新鲜的菌落接种于2ml-5mlSOB培养基的试管中,37℃振荡培养过夜,转速控制在180rpm。2)第二天,取过夜培养物250uL转接入含50ml SOB培养基(1 ...

