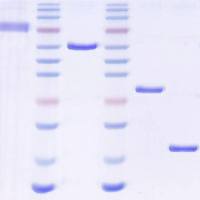
GPI蛋白
互联网
GPI蛋白
本文提供了有关糖基磷脂酰肌醇(GPI,是许多蛋白以及病毒蛋白结合于膜上的锚点)的信息。同时,网站还提供了许多相关链接和资源。
GPI overview
GPI proteins in chickens
GPI proteins in Drosophila
GPI in prion proteins
Human GPI proteins: protectin (CD59) and homologous restriction factor (HRF, C8bp, MIP)
GPI proteins in yeast (14 in whole genome
Prion oligosaccharide abstracts
Prion proteins from all species sequenced so far (from human to chicken) contain a C-terminal GPI-anchor, a post-translational modification that replaces an initial hydrophobic terminus with glycosyl phophotidylinositol, serving to attach the mature protein to the outside of the cytoplasmic membrane, while allowing for later release by phospholipase. It seems likely that salmon prion will contain this modification, pushing the date of origin of this modification of prion protein back to at least 440 million years ago.
GPI-anchors are not common as a protein modification. In yeast, where all GPI proteins are known, only 15 of the 5,790 (so 1 in 400) proteins have GPI-anchors and even these have considerable homology with each other. When mammalian homologues of yeast GPI proteins are known, these have the GPI-modification as well. In such protein families, the modification has evidently been in continuous use for a billion years or more. Yeast GPI biosynthetic enzymes are likewise homologous to mammals counterparts.
GPI proteins have been found in a wide variety of eucaryotes : mammals (45 in humans), chickens (10), fish, rays, sea urchin, fruit flies (5), silk moth, ticks, grasshopper, protozoa (trypanosomes, leishmanii, paramecium), fungi, slime mold, unicellular green alga, mung bean, even herpes virus (simian surface glycoprotein), but not in bacteria, and oddly nothing reported from nematode (out of 1208 proteins).
GPI-proteins are evenly split between enzymes and binding, recognition, and transport non-catalytic proteins. This split correlates fairly strongly with whether internal tandem repeats are present (non-catalytic) or not (catalytic). A GPI-anchor unsurprisingly implies a signal peptide but by no means conversely. There is a division between O- and N-glycosylation. Identified functions are clearly appropriate to the extra-cytoplasmic location; GPI proteins are over-represented in neurons.
Human genetic disorders have been identified in five GPI proteins other than prion These include Marfan's syndrome [FBN1 gene] and Charcot-Leyden crystals [LPPL gene], alkaline phosphatase: infantile hypophosphatasia, lipoprotein lipase: chylomicronemia syndrome, and glypican-3: Simpson-Golabi-Behmel syndrome> A defect in a GPI biosynthetic enzme causes paroxysmal nocturnal hemoglobinuria.
The question arises, how ancient is the GPI-anchor in prion protein, how often and which sorts of new proteins are recruited to GPI-status, how many super-families of GPI proteins exist and to which does prion protein belong, what sort of internal structural motifs occur, where are close but non-GPIhomologues located in the cell, is prion protein more likely catalytic than structural, and finally, does analysis of GPI-proteins help identify the normal function of prion protein or suggest other candidates for prionic disease?
<center> </center>
Review of GPI-anchored membrane proteins
Brown D; Waneck GL J Am Soc Nephrol 3: 895-906 (1992)
Many proteins of eukaryotic cells are anchored to membranes by covalent linkage to glycosyl-phosphatidylinositol (GPI). These proteins lack a transmembrane domain, have no cytoplasmic tail, and are, therefore, located exclusively on the extracellular side of the plasma membrane.
GPI-anchored proteins form a diverse family of molecules that includes membrane-associated enzymes, adhesion molecules, activation antigens, differentiation markers, protozoan coat components, and other miscellaneous glycoproteins. In the kidney, several GPI-anchored proteins have been identified, including uromodulin (Tamm-Horsfall glycoprotein), carbonic anhydrase type IV, alkaline phosphatase, Thy-1, BP-3, aminopeptidase P, and dipeptidylpeptidase.
GPI-anchored proteins can be released from membranes with specific phospholipases and can be recovered from the detergent-insoluble pellet after Triton X-114 treatment of membranes. All GPI-anchored proteins are initially synthesized with a transmembrane anchor, but after translocation across the membrane of the endoplasmic reticulum, the ecto-domain of the protein is cleaved and covalently linked to a preformed GPI anchor by a specific transamidase enzyme.
At least one human disease, paroxysmal nocturnal hemoglobinuria, is a result of defective GPI anchor addition to plasma membrane proteins. Although it remains obscure why so many proteins are endowed with a GPI anchor, the presence of a GPI anchor does confer some functional characteristics to proteins:
(1) it is a strong apical targeting signal in polarized epithelial cells;
(2) GPI-anchored proteins do not cluster into clathrin-coated pits but instead are concentrated into specialized lipid domains in the membrane, including so-called smooth pinocytotic vesicles, or caveoli;
(3) GPI-anchored proteins can act as activation antigens in the immune system;
(4) when the GPI anchor is cleaved by PI-phospholipase C or PI-phospholipase D, second messengers for signal transduction may be generated;
(5) the GPI anchor can modulate antigen presentation by major histocompatibility complex molecules.
Identifying Lipid Anchors:
The protein modification reactions which bind lipid molecules to proteins are important because a linked lipid moiety can be integrated into various membranes and can anchor the bound protein. All proteins linked to the glycosyl-phosphatidylinositol (GPI) molecules are thought to be anchored at the extracellular surface of the plasma membrane. PSORT recognizes GPI-anchored proteins by the knowledge that most of them are predicted to be type Ia membrane proteins with very short cytoplasmic tail (within 10 residues) and uses the result for the prediction of the localization site (plasma membrane) of the modified protein.
<center> <p> Twelve chicken GPI-anchor proteins:</p></center>
AXO1_CHICK Axonin-1 precursor: 1036 aa P28685
EPL6_CHICK EPH-related receptor tyrosine kinase ligand 6 precursor (lerk-6)200 aa P52802
G55A_CHICK Neurite inhibitor gp55a (fragment) 274 aa Q98892
GLYP_CHICK Glypican-1 precursor (heparan sulfate proteoglycan core protein). P50593 550 AA.
LAMP_CHICK Limbic system-associated membrane protein precursor (e19s). Q98919 338 AA.
CADD_CHICK T-Cadherin precursor (truncated-cadherin) (cadherin-13).712 AA. P33150
CNTR_CHICK Ciliary neurotrophic factor recepto r alpha precursor P51641 362 AA
LIPL_CHICK Lipoprotein lipase precursor (EC 3.1.1.34) (LPL) P11602 490 AA
TECB_CHICK Beta-tectorin precursor 329 AA P54097
CONT_CHICK Contactin precursor (neural cell recognition molecule f11).1010 AA. P14781
THY1_CHICK THY-1 Membrane glycoprotein precursor (thy-1 antigen) 160 AA. Q07212
<center> </center>
AXO1_CHICK Axonin-1 precursor: 1036 AA P28685
Axon-associated cell adhesion molecule (axcam) which promotes neurite outgrowth by interaction with the axcam l1 (g4) of neuritic membrane. Atached to the neuronal membrane by agpi-anchor. Belongs to the immunoglobulin superfamily. contains six c2-like domains followed by four fibronectin type iii-like domains
signal 1 23 or 25 (potential).
chain 24 1036 axonin-1.
propep ? 1036 removed in mature form.
domain 49 113 ig-like c2-type domain.
domain 143 211 ig-like c2-type domain.
domain 249 308 ig-like c2-type domain.
domain 336 397 ig-like c2-type domain.
domain 428 490 ig-like c2-type domain.
domain 518 589 ig-like c2-type domain.
domain 599 608 hinge (potential).
domain 601 607 gly/pro-rich.
domain 608 709 fibronectin type-iii.
domain 710 811 fibronectin type-iii.
domain 812 912 fibronectin type-iii.
domain 913 1009 fibronectin type-iii.
mod_res ?24 ?24 blocked.
carbohyd 71 71 potential.
carbohyd 199 199 potential.
carbohyd 456 456 potential.
carbohyd 472 472 potential.
carbohyd 493 493 potential.
carbohyd 520 520 potential.
carbohyd 770 770 potential.
carbohyd 900 900 potential.
carbohyd 914 914 potential.
<center> </center>
EPL6_CHICK EPH-related receptor tyrosine kinase ligand 6 precursor (lerk-6)200 aa P52802
Binds to the receptor tyrosine kinases cek7, mek4 and sek.
Atached to the membrane by a gpi-anchor
Homologous to oher members of the eplg family.
signal 1 22
ligand 6.
carbohyd 36 36
carbohyd 161 161
carbohyd 175 175
<center> </center>
G55A_Chick Neurite inhibitor gp55a (fragment) 274 aa. q98892
Inhibits neurite outgrowth.
Atached to the membrane by a gpi-anchorsimilarity: belongs to the immunoglobulin superfamily. Contains three c2-like domains.
Belongs to the obcam subfamily, immunoglobulin fold
non_ter 1 1
domain<1 52 ig-like c2-type domain.
domain 80 138 ig-like c2-type domain.
domain 166 232 ig-like c2-type domain.
disulfid 87 131
disulfid 173 225
carbohyd 70 70
carbohyd 214 214
carbohyd 222 222
carbohyd 235 235
<center> </center>
GLYP_CHICK Glypican-1 precursor (heparan sulfate proteoglycan core protein). P50593 550 AA.
Cell surface proteoglycan that bears heparan sulfate.
Attached to the membrane by a gpi-anchor
Belongs to the glypican family.
signal 1 20
chain 21 ? glypican-1.
propep ? 550 removed in mature form (potential).
carbohyd 76 76
carbohyd 113 113
carbohyd 382 382
carbohyd 52 52 glycosaminoglycan (potential).
carbohyd 483 483 glycosaminoglycan (potential).
carbohyd 485 485 glycosaminoglycan (potential).
carbohyd 487 487 glycosaminoglycan (potential).
<center> </center>
LAMP_CHICK Limbic system-associated membrane protein precursor (e19s). Q98919; 338 AA.
Mediates selective neuronal growth and axon targeting. Probably serves as a recognition molecule for the formation of limbic connections
Attached to the membrane by a gpi-anchorsimilarity:
Belongs to the immunoglobulin superfamily. contains three c2-like domains. belongs to the obcam subfamily.
signal 1 28
chain 29 ? limbic system-associated membraneprotein.
propep ? 338 removed in mature form (potential).
domain 46 118 ig-like c2-type domain.
domain 146 204 ig-like c2-type domain.
domain 232 297 ig-like c2-type domain.
disulfid 53 111
disulfid 153 197
disulfid 239 290
carbohyd 40 40
carbohyd 66 66
carbohyd 136 136
carbohyd 148 148
carbohyd 279 279
carbohyd 287 287
carbohyd 300 300
carbohyd 315 315
<center> </center>
LIPL_CHICK Lipoprotein lipase precursor (EC 3.1.1.34) (LPL) P11602 490 AA
The primary function of this lipase is the hydrolysis
of triglycerides of circulating chylomicrons and very low density
lipoproteins (vldl). The enzyme functions in the presence of
apolipoprotein c-2 on the luminal surface of vascular endothelium.
catalytic activity: triacylglycerol + h(2)o = diacylglycerol +
a fatty acid anion.
Subunit: homodimer, interact with apolipoprotein c-2.
Partial homology with other lipases (pancreatic, gastric, hepatic, lingual, lipoprotein, bacterial, etc.).
signal 1 25
chain 26 490 lipoprotein lipase.
act_site 159 159 charge relay system (by similarity).
act_site 183 183 charge relay system (by similarity).
act_site 268 268 charge relay system (by similarity).
domain 319 331 heparin-binding (potential).
disulfid 54 67
disulfid 243 266
disulfid 291 310
disulfid 302 305
disulfid 445 465
carbohyd 70 70
carbohyd 386 386
<center> </center>
TECB_CHICK Beta-tectorin precursor. 329 AA P54097
Extracellular matrix. The n-terminal is blocked. n-glycosylated.
Tissue specificity: inner ear (basilar papilla, clear cells and lagena macula).
Contains a zp domain, which currently has been found in zp2, zp3, gp2, tgfr-3 and uromodulin.
signal 1 17
chain 18 329 beta-tectorin.
domain 91 283 zp (br> carbohyd 80 80
carbohyd 104 104
carbohyd 116 116
carbohyd 145 145
<center> </center>
CONT_CHICK Contactin precursor (neural cell recognition molecule f11).1010 AA. P14781
Mediates cell surface interactions during nervous system development.
Belongs to the immunoglobulin superfamily. contains six c2-like domains followed by four fibronectin type iii-like domains.
signal 1 19
chain 20 ? contactin.
propep ? 1010 removed in mature form.
domain 50 113 ig-like c2-type domain.
domain 143 210 ig-like c2-type domain.
domain 247 308 ig-like c2-type domain.
domain 336 389 ig-like c2-type domain.
domain 420 482 ig-like c2-type domain.
domain 510 581 ig-like c2-type domain.
domain 593 599 gly/pro-rich.
domain 600 701 fibronectin type-iii.
domain 702 803 fibronectin type-iii.
domain 804 900 fibronectin type-iii.
domain 901 996 fibronectin type-iii.
carbohyd 200 200
carbohyd 249 249
carbohyd 329 329
carbohyd 448 448
carbohyd 464 464
carbohyd 485 485
carbohyd 512 512
carbohyd 582 582
carbohyd 924 924
<center> </center>
CADD_CHICK T-Cadherin precursor (truncated-cadherin) (cadherin-13).712 AA. P33150
Tissue specificity: neural tissues. also found in muscles; kidney and retina.
Similarity: strong to other cadherins.
signal 1 22
propep 23 138
chain 139 693 t-cadherin.
propep 694 712 removed in mature form
repeat 139 245 cadherin 1.
repeat 246 363 cadherin 2.
repeat 364 477 cadherin 3.
repeat 478 583 cadherin 4.
repeat 583 693 cadherin 5.
carbohyd 86 86
carbohyd 382 382
carbohyd 500 500
carbohyd 530 530
carbohyd 638 638
carbohyd 671 671
lipid 693 693 gpi-anchor
Cadherins are a family of animal glycoproteins responsible for calcium-
dependent cell-cell adhesion. Cadherins preferentially interact with
themselves in a homophilic manner in connecting cells; thus acting as both
receptor and ligand. A wide number of tissue-specific forms of cadherins are
known:
- Epithelial (E-cadherin) (also known as uvomorulin or L-CAM) (CDH1).
- Neural (N-cadherin) (CDH2).
- Placental (P-cadherin) (CDH3).
- Retinal (R-cadherin) (CDH4).
- Vascular endothelial (VE-cadherin) (CDH5).
- Kidney (K-cadherin) (CDH6).
- Cadherin-8 (CDH8).
- Osteoblast (OB-cadherin) (CDH11).
- Brain (BR-cadherin) (CDH12).
- T-cadherin (truncated cadherin) (CDH13).
- Muscle (M-cadherin) (CDH14).
- Liver-intestine (LI-cadherin).
- EP-cadherin.