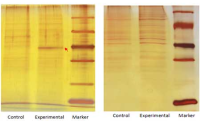
病原体逃逸抗感染免疫的机制病毒因素
互联网
2283
病原体逃逸抗感染免疫的机制病毒因素
病原体可利用各种机制逃逸机体的免疫防御作用。由于病原体导致机体感染引起疾病是病原体和宿主机体相互作用的结果。因此,机体病原体逃逸抗感染免疫的机制主要涉及病原体本身的因素和机体的因素。
不同类型的病原体逃逸机体的抗感染免疫的机制不尽相同,但主要可归纳于隐匿、诱变和抑制。隐匿是指病原体通过感染免疫细胞本身或寄生于免疫应答不易到达之处而逃逸免疫攻击;诱变是指病原体通过改变自身的抗原特性,从而逃逸业已诱生的免疫应答的作用;抑制是指病原体通过其结构和非结构产物,拮抗、阻断和抑制机体的免疫应答。
可编码多种蛋白从不同水平干扰宿主的抗感染免疫防御机制。例如,
①抗原变异:流感病毒的包膜蛋白血凝素和神经氨酸酶构成主要抗原和中和抗原,但可经常地持续性地发生突变,逃逸已建立的抗感染免疫抗体的中和和阻断作用,造成多次流感世界大流行和连续不断的地区性小流行。小幅度的抗原变异称为抗原漂移(antigenicdhk),大幅度的抗原变异称为抗原转换(antigenicshift)。HIV病毒的包膜蛋白也高度突变,其突变速度要比流感病毒快65倍左右。
②抑制被感染细胞的凋亡:被感染细胞的自发凋亡也是固有免疫防御的策略之一。许多痘病毒编码丝氨酸蛋白酶抑制剂(serpin),抑制凋亡途径中caspase酶的激活,从而抑制凋亡。而SV40、腺病毒、HBV等表达的大T抗原、E1B―55K、pX基因产物,可结合P53并抑制其功能,抑制宿主细胞凋亡。而疱疹病毒则编码Bcb2类似物,抑制凋亡。
③阻断补体激活:痘苗病毒产生能与CAb结合的蛋白,阻抑补体经典途径的激活。单纯疱疹病毒的一种糖蛋白,与C3b结合后可同时抑制补体经典和替代激活途径。
④抑制NK细胞的抗病毒杀伤功能:已知NK细胞是抗病毒感染的第一道固有免疫防线。HIV具有显著的抗NK细胞杀伤功能。已知人类正常细胞表面的HLA-C和HLA-E与NK细胞的抑制受体结合可阻止杀伤;而HIV选择性下调HLA-A和HLA-B分子表达,这两类分子正是CTL识别的I类分子,因此,HIV不仅逃避NK细胞的天然杀伤,同时逃避特异性CTL的识别杀伤。CMV也有抑制NK细胞杀伤功能。
⑤编码细胞因子和趋化因子的类似物,干扰其重要调节功能:如EB病毒产生IL-10的类似物BCRF-1,抑制Thl细胞产生IL-2、TNF-α和IFN β。EB病毒、HSV、腺病毒、HBV分别编码I型IFN的类似物EBNA-2、γ134.5、E1A和末端蛋白,阻断或下调IFN的抗病毒功能。已知PKR(双链RNA依赖的蛋白激酶)通路参与IFN合成、对于清除病毒感染十分重要,因此许多病毒和EB病毒、HSV、HCV、HIV编码PKR的抑制剂、竞争结合物或裂解酶,终止这一通路从而抑制对病毒的清除。多种痘病毒编码针对IFN β、IFN-α/β、ILl、IL-6、IL-8、TNF的分泌型受体,从而中和上述细胞因子的抗病毒活性以及重要免疫调节活性。许多γ/β疱疹病毒分泌细胞因子或趋化因子的类似物,称为病毒因子(virokine),如HHV-8和EB病毒均感染B细胞、又分别产生IL-6和ILl0的类似物,均促进B增殖和应答,扩大感染宿主细胞数目,同时抑制CTL的诱导。
⑥干扰抗原多肽的I类提呈、抑制CTL诱生:腺病毒、HIV等分别合成E1A、Tat蛋白,抑制MHCI类分子转录,HCMV则编码US2/USll、ULl8蛋白,分别降解MHC工分子和竞争结合β2m分子。腺病毒和HSV、HCMV还编码病毒蛋白阻断TAP通道蛋白,使MHCI~抗原提呈途径受阻。最终无法有效诱导抗病毒CTL应答。
各种病毒综合应用多种机制逃逸并破坏免疫系统。在HIV感染中,HIV糖蛋白gpl20可脱落,与CD4分子结合并封闭CD4分子,使其免疫功能丧失;HIV的Tat蛋白可促进FasL的表达,抑制凋亡抑制蛋白Bcl 2的表达,从而促进CD4T细胞凋亡;HIV的Vpr和gpl60也可诱导CD4 T和CD8 T凋亡,使数量减少;在HBV慢性感染过程中,HBV改变了B细胞应答特性、CTL应答特性和APC抗原提呈功能,来逃避免疫攻击。慢性乙肝患者往往抗HBcAg/HBeAg特异性CD4T应答下调或缺失,导致HBcAg特异性CTL数量下调或消失,而这一应答对于清除HBV感染细胞非常关键。此外,抗HBV次要CTI。表位(HBsAg内表位)的CD8CTL虽然存在,但其TCR表型发生改变,使不能被MHO多肽四聚物技术检测,这种TCR表型改变的特征在其他病毒持续感染中也可见到。抗HBsAg特异性抗体应答同样在慢性乙肝患者中下调。