材料与仪器
步骤
增殖分化调控机制的研究
载 体 的 制 备
目的基因的克隆是分子生物学实验中的核心内容,起着承上启下的作用,通过克隆技术可实现目的基因的无限扩繁、长期保存的目的;利用该技术能实现基因-组、基因改造、构建工程菌和转基因动植物。基因克隆技术主要包括载体的获取、酶切、脱憐、连接和转化及筛选、鉴定等内容 (奥斯伯等 2001)。在干细胞增殖、分化调控机制的研究中常用到的克隆载体主要有以下两类:①宿主为大肠杆菌的细菌质粒;②动物细胞为宿主的动物病毒和以动物病毒为基础改造的穿梭质粒。
载体特点:①在宿主细胞中具独立复制能力;②带抗性等选择标记;③有合适的限制性内切核酸酶位点,可插人一定片段长度的外源 DNA 而不影响自身复制。
质 粒 提 取
质 粒 D N A 的提取方法比较常见,基本步骤多为三步:①细菌培养和质粒的扩增;②细菌菌体的裂解;③ 质 粒 DNA 的纯化。现在常用的是质粒提取试剂盒。该方法方快 捷 ,具体步骤可参考其使用手册。
注意:在 质 粒 DNA 提取中,质 粒 D N A 的质量和产量在很大程度上受培养条件的影响 ,通过一些实验发现,下列培养方式能获得较好的结果。①培养应用菌龄不超过 4 天的平板单菌落作为起始菌。②培养菌应在 37℃、 220r/min 下 生 长 16—— 18 h。③不能用经储藏的培养菌直接进行质粒提取制备。
D N A 片 段 的 酶 切
限制性内切核酸酶 (restriction endonuclease) 是一类能识别双链 DNA 分子中特异核苷酸序列的 DNA 水解酶,是体外剪切基因片段的重要工具。
1 ) 主要设备与试剂
离心机、恒温水浴、电泳仪、电泳槽、质粒、 TAE 缓冲液、限制性内切核酸酶
2 ) 操作步骤
用于质粒的酶切鉴定,一 般 是 20ul体积中含有0.2〜Iug D N A , 酶切反应体系为:
质 粒 DNA 10ul
缓冲液 (IOx) 2ul(双酶切时要选取两种酶都适合的缓冲液)
限制性内切核酸酶 1ul(双酶切时两种酶各加 1 ul)
双蒸水 7ul
⑴ 在 E p 管中加入 7 ul双蒸水。
⑵ 加 人 IOx 缓 冲 液 2ul 。
(3) 加 限 制 酶 1 ul。
⑷加入质粒 10 ul
(5) 离 心 ,混匀。
(6) 37℃水浴 l——2 h。
(7) 取 反 应 液 ,电泳鉴定。
3 ) 注意事项
⑴ 样 品 加 入 次 序 为 水 、缓冲液、 D N A ,最后为酶,不应颠倒。
(2) 加酶步骤要在冰浴中进行,在加酶前应先将水、缓冲液及待切 D N A 混匀。
(3) 反应体系的体积要尽量少 , 一 般 水 解 0.2〜 Iug D N A 时 ,应将体积控制在 20——30 ul内。保证限制酶活性不受抑制。
⑷为了控制反应体积和促进反应进行,要求模板 D N A 的浓度应很高,如 底 物 D N A浓度过低则应进行浓缩。
(5) 本实验中双酶切时,因两种酶的缓冲液盐浓度要求相同,所 以 ,可在同—反应体系中完成双酶切。但如进行双酶切时,两种酶所需缓冲液盐浓度要求不同,则不能将两种酶同时加入同一体系进行反应,此时可采取一种酶水解完后,进行有机溶剂抽提,沉淀、干燥后再复溶,然后进行第二种酶切。
D N A 片 段 回 收
回收是指从电泳介质中纯化出目的片段。在试验中待回收的片段主要有酶切片段和各 种 P C R 片段;回收的介质主要为琼脂糖;回收的方法主要有微量柱回收、玻璃奶回收。这里以玻璃奶法为例介绍 D N A 片段回收的方法。
1 ) 主要设备与试剂
离心机、烘箱、恒温水浴、玻璃奶、无水乙醇、溶胶液、漂洗液。
2 ) 操作步骤
(1) 从琼脂糖凝胶上切下目的条带放入 1.5 ml E p 管中,用 一 个 Tip 头捣碎。
⑵ 准 确 估 计 凝 胶 的 体 积 ,加 入 3 倍体积的溶胶液,室温放置 5m i n 或 50℃:保 温 3 min,其间轻摇 E p 管几次使胶完全溶化
。
⑶ 向 溶 胶 中 加 入 IOpl 玻璃奶,颠倒混勻,冰 浴 lOmin,并且间隔 2〜 3m i n 颠倒混匀一 次 , 12 000r/m i n 离 心 30s,吸弃上清。
(4) 将浓缩漂洗液按 3 : 7 与无水乙醇配成工作液,然后向玻璃奶中加入 2 5 0 ul ,用加样器吸漂洗液轻柔地将玻璃奶冲散混匀, 12 OOOr/m in 离 心 30s ,吸弃上清。
(5) 重复第 (4) 步一次。吸取完漂洗液后再离心 10s,用 Tip 头将最后一点儿漂洗液吸干 净 ,然后放置于 37℃ 温箱中干燥 20 min。
(6) 加 20ul 无菌水,用 Tip 头轻轻混勻, 60T ;水 浴 5 min, 12 OOOr/m i n 离 心 lmin,回收上清即为纯化的 P C R 片段。 -20℃ 储存。
3 ) 注意事项
(1) 玻璃奶使用之前一定要融化并充分混匀。
(2) 干燥后加无菌水后要充分吹打。
基 因 的 重 组 连 接
重组连接是指将目的基因用 D N A 连接酶连接在合适的质粒载体上,这是基因重组、基因改造的重要中间环节。连接反应可在溶液中进行也可在琼脂块中进行。在本部分仅以重组克隆为目的介绍相应的连接类型。在基因重组克隆中,外 源 D N A 与载体的连接是承上启下的一步操作,根据连接片段末端类型不同,连接方式有黏端连接、平端连接、平黏端连接及单碱基配对的 T 载体连接等。
主 要 设 备 与 试 剂
离心机、恒温设备、 T 4 D N A 连接酶、 10xT4 D N A 连接酶缓冲液。
操 作 步 骤
连接
1) 取干净、灭菌 E p 管,加入下列各试剂
T4 DNA 连接酶缓冲液 1ul
载 体 DNA 片段 1ng
插入目的片段 xng
连接酶 IU(平末端,如为黏端连接,则酶量可小于 1u)
加无菌水至 IOul
X = 目的片段与载体片段的分子比例 X 载 体 n g 数 X 目的片段的碱基数/载 体 n g 数
(目的片段/载体片段的分子比例一般为 3 : 1)
2) 连接反应
2 2 ℃ 保 温 3 h 或 4℃过 夜(Promega 连接酶,黏端连接);
或 22〜 2 6 ℃保 温 Ih (B R L 连接酶,黏端连接);
或 22℃保 温 4—— 16 h (Promega 连接酶,平端连接);
或 14℃ 保 温 16——24 h (B R L 连接酶)。
3 ) 注意事项
(1) 分子生物学实验中主要采用 T 4 D N A 连接酶。
(2) 在分子生物学实验中连接酶主要用于:①基因重组中载体与外源基因的连接;② 接 头 与 D N A 片段的连接,这种连接一般发生在文库构建、分子标记的 A F L P 及 差 ^表达的研究中等。
(3) 其中黏端连接的效率最高,黏端连接与平黏连接中外源 D N A 片段在载体中的插入有方向性。平端连接的效率最低,且载体发生自连的比例较髙,因此在克隆操作中为了克服平连的弊端,通常可通过:① T d T 酶在载体和外源 D N A 片段上转移一互补序列 (核苷酸投影法);②在载体和外源 D N A 片段上加上人工接头,变平连为黏连。
连接产物的转化
通过转化可以实现大量扩增质粒。转化的方法有多种,如电转化法、 CaCl2 法等。一般使用 CaCl2 法 ,从而改变细胞膜的结构,使 外 源 D N A 可以穿过细胞膜进入细胞,然后在选择性培养基中培养转化处理过的细菌,转化成功的细菌可以在抗性培养基上生长形成菌落。
1 ) 主要设备与试剂
摇床、离心机、恒温设备、 L B 培养液、 CaCl2(0.1mol/L )。
2 ) 操作步骤
(1) 从新鲜的平板上挑取一个 D H 5α 单克隆, 37℃摇振培养过夜。
(2) 按 I : 100 的比例将过夜培养的菌液加入新鲜的 L B 培养液中,于 37℃继续培养至 OD600为 0.3——0.4。
(3) 将菌液转移到预冷的 50m l 离心管中, 4 ℃ 、: IOOOg 离 心 15m i n 回收菌体。
(4) 倒出上清,将 管 倒 置 Imin 以使最后残留的痕量液体流尽。
(5) 用 IOml 冰预冷的 0.1mol/L CaCl2 重悬每份沉淀,同上离心收集菌体。
(6) 重复步骤⑷。
(7) 每 50m l 初始培养物用 2m l 冰预冷的 0.1mol/L CaCl2 重悬,此即感受态细菌。
(8) 取 200 叫感受态细菌和 D N A (≤10ul, ≤ 5 0 ng 伽入到一个冰冷的聚丙烯管中,轻轻旋转混匀,冰上放置 30 min。
(9) 将管放到 42 ℃水浴中,恰恰放置 90s,其间不要摇动。
(10) 快速将管转移到冰浴中,冷 却 l——2 min。
(11) 每 管 加 800ul L B 培养液, 37 ℃温和摇振培养 60m i n 使细菌复苏,并且表达质粒编码的抗生素抗性标记基因。
(12) 取 2 0 0 0 菌液涂在含 A m p 或 K a n 平板上,将平板置于室温直至液体被吸收,然 后 于 3 7 ℃ 倒置培养过夜。
3 ) 注意事项
(1) 热激温度和时间要严格控制好。
(2) 含 A m p 琼脂平板培养时间不应超过 16 h ,否则容易产生卫星菌落。
⑶ 如 果 是 与 T 载体的连接产物,则将菌液涂布在含 X -gal 和 I P T G 的 A m p 或 Kan平板上。
病毒载体的制备 (详见细胞转染部分)
1 ) 逆转录病毒载体的构建
⑴ 改 造 病 毒 基 因 组 ,切 除 野 生 型 的 娜 、 poZ、 e«v 结构基因,换为目的基因,保留包装信号和 L T R S 序列。
⑵将重组的病毒载体导入包装细胞,包装出有感染能力的逆转录病毒颗粒。
(3) 从包装细胞培养上清中获得包装好的缺陷型病毒,用它转导带有该病毒受体的祀细胞,使目的基因稳定整合至靶细胞染色体。
注意:①插入的目的基因不能超过 7kb , 否则逆转录病毒载体将不能有效包装。②外源基因导入靶细胞的效率很高,通 常 Im l 含病毒的培养基可感染 1 〇 6个觀细胞。
2 ) 腺病毒载体的构建 ( ;这部分已经商品化,不做详细介绍)
(1) 构建含外源目的基因的腺病毒穿梭载体。
(2) 制备重组质粒。
(3) 转 染 HEK293 细胞。
(4) 收获含重组腺病毒的上清进行滴度测定和纯化。
(5) 转染靶细胞。
注意: 一般插入外源 DNA 长度不得超过 2kb, 但是通过缺失部分病毒基因组可扩大外源 D N A 的插入容量。
蛋 白 质 的 制 备
在干细胞增殖、分化调控机制的研究中,常需要进行目的蛋白质的表达。首先我们通过载体制备将目的基因插入表达载体,然后再将该重组载体导入活细胞,以大量表达所需目的蛋白质。
外源基因可以在原核表达,也可以在真核细胞中进行表达。外源基因的原核表达是目的基因在原核细胞中以发酵的方式快速、高效的合成基因产物,常用于原核表达的细胞是大肠杆菌。其特点是:操作简便,而且在较短时间可以实现蛋白质的大量表达,因
此成为科研应用中蛋白质表达的主要宿主。但是大肠杆菌进行蛋白质表达也存在一些缺陷 :①真核蛋白在大肠杆菌中的表达没有得到正确的修饰;②在大肠杆菌中表达蛋白时常形成沉淀成为包含体。
诱导
常用表达用宿主菌株为 DE3 BL2 1 菌 ,先要准备合适的感受态细胞,取 质 粒 Ing 用无菌水稀释 5 0 倍 ,按前述转化步骤进行操作,筛选出阳性克隆后提质粒并经 D N A 序列测 定 ,确定无误后进行如下操作。
1 ) 主要设备与试剂
摇床、 SDS-PAGE 电泳仪、离心机、 LB 培养基、 IPTG、 Tris • HCI(20mmol/L pH8)、2xloading buffer
2 ) 操作步骤
⑴从新鲜的划线平板中挑取单克隆,接至一定体积的含合适抗生素的 L B 中, 37℃摇床振摇培养至OD600为 0.4〜 0.6。
(2) 如果表达载体的原核启动子为 PL 启动子,则 在 30〜 32℃ 培养数小时,使培养液的 OD600达 0.4〜 0.6, 迅速使温度升至 4 2 ℃ 继 续 培 养 3——5h。如果表达载体的原核启动子为 ta c 等 ,则 37T 培养细菌数小时达到对数生长期后加 IPTG至终浓度为lmmol/L,继续 培 养 3——5 h。
(3) 取 出 Im l菌液做分析用, 1 〇〇〇g发离心 lmin,沉 淀 加 l 〇〇ul 2xloadingbuffer, IOO℃加 热 3 min, 上 样 进 行 SDS-PAGE。
⑷ 将 摇 瓶 置 于 冰 上 5 min。4℃ 、 5 000g 离 心 5m in 收集菌体,重悬细胞于〇.2 5 倍体积预冷的2 〇 mol/L Tris HCl(pH 8.0) 中,离心。菌体保存于-7〇℃冻存以备分离基因表达产物所用。
3 ) 注意事项
不同的表达质粒,因启动子不同,诱导表达方法并不完全相同,可根据具体情况而定。
细 菌 的 裂 解
细胞破碎是提取胞内产物的关键性步骤。常用的研究方法有酶溶法和超声破碎法。
溶法
酶溶法就是用生物酶将细胞壁和细胞膜消化溶解的方法。常用的溶解酶有溶菌酶(Iysozyme)
1 ) 主要设备与试剂
离心机,裂解缓冲液 (lmmol/L EDTA, 100 mmol/L NaCl,50 mmol/L Tris • Cl,p H 8 . 0),蛋白酶抑制剂 (50 mmol/L ) , 溶菌酶(10m g /ml),脱氧胆酸, D N a SeI(lmg/ml)。
2 ) 操作步骤
(1) 4℃ 、 500 g 离 心 15 min,收集诱导表达的细菌(1000 ml)。
(2) 弃上清液,每克菌加3m l 裂解缓冲液,悬浮沉淀。
(3)每 克 菌 加 3ul 5Ommol/L 的 蛋 白 酶 抑 制 剂 苯 甲 磺 酰 氟 (phenylmethyIsulfonylfluoride, PMSF), 80ul 溶菌酶(10m g /ml),搅拌 20 min。
(4) 在搅拌下,每克菌加 4m g 脱氧胆酸。
(5) 溶 液 变 得 黏 稠 时 每 克 菌 加 20ul DNaseI(lmg/ml)。
(6)室温放置至溶液不再黏稠。
3 ) 注意事项
步骤 (2)〜 (4) 在冷室中进行。
超声破碎法
声 频 高 于 15〜 20kHz的超声波在高强度声能输入下可以进行细胞破碎,在处理少量样品时操作简便,液体量损失少,同时还可对染色体 D N A 进行剪切,大大地降低了液体的黏度,有利于后续目标产物的分离。
1 ) 主要设备与试剂
离心机、 SIDS-PAGE 电泳仪、 T E buffer、 2xloading buffer。
2 ) 操作步骤
⑴收集诱导表达的细菌(IOOOml),4℃ 、 500 g 离 心 15 min。
⑵ 弃 上 清 液 ,每克菌加3 ml T E buffer。
(3) 根据厂家提供的超声波仪的数据进行破菌。
⑷ 10 000g 离 心 15 min, 分别收集上清液和沉淀。
(5) 取 少 量 上 清 液 和 沉 淀 加 入 等 体 积 的 2xloading b u ffe r, 进 行 SDS-PAGE电泳。
3 ) 注意事项
(1) 超声破碎与声频、处理时间、细胞浓度、菌种类型等许多因素有关,实验时要根据具体情况掌握。
(2) 超声波破菌前,细胞经数次冻溶后更容易破碎。
包 含 体 的 分 离 、 溶 解 和 复 性
在以大肠杆菌为宿主菌高效表达外源基因时,表达蛋白常常在细胞质内聚集,形成包含体 (inclusion body)。包含体形成后,表达蛋白不具有物理活性,因此必须溶解包含体对表达蛋白进行复性。由于包含体是致密的,易于沉淀,所以能用脱氧胆酸盐洗涤得到纯度较高的制备物。包含体离心沉淀后,可用尿素等洗涤。为获得可溶性的活性蛋白,必须要将洗涤过的包含体溶解,然后进行重折叠,最后用离子交换或亲和层析的方法对重折叠后的蛋白质进行精制。
分离
1 ) 主要设备与试剂
离心机、 SDS-PAGE 电泳仪、尿素、 Tris . ClCaimolZL, p H 8.5)、 2xloading buffer。
2 ) 操作步骤
(1)4℃,12 OOOg离心细菌裂解液 15 min。
(2) 弃上清液,每 克 菌 加 Iml 无菌水使悬浮,分 别 取 1 0 0 ul 分 装 4 个 管 ,其余备用。
⑶ 离 心 同 步 骤 (1)。
(4) 弃上清液,用 lOOul O.lmolTLTris • Cl(p H 8.5),含不同浓度尿素(0.5mol/L 、lmol/L )的溶液悬浮沉淀。
(5)离 心 。
(6) 分别吸出并保留上清液,用 100ul H2O 重新悬浮沉淀。
(7) 分别取10 叫上清液和重新悬浮的沉淀,加 10ul 2xloading buffer进 行 SDS-PAGE电泳。
包含体的溶解
1 ) 主要设备与试剂
SDS-PAGE 电泳仪,裂解缓冲液 (含 0.1 m m 〇 l/L P M S F ,8mol/L 尿 素 ,10 mmol/L D T T ),K H 2P O 4(50 mmoiyL, p H 10.7), E D T A (lmmol/L , p H 8.0), NaCl(50 mmol/L )
2 ) 操作步骤
(1) 用 IOOul裂解缓冲液溶解包含体。
(2) 室 温 放 置 1h
(3) 加 9 倍体积的 P A G E 电泳缓冲液,室温放置30 min, 用 K O H 保 持 p H 为 10.7。
⑷ 用 H C l 调 至 p H 8.0,最少在室温放置30 min。
(5) 室温、 10 000g离心15min。
(6) 吸出上清液并保存,用 100 ul IxSDS-Ioading buffer重新溶解沉淀。
(7) 取 10 ul 上 清 液 ,加 10ul 2xloading buffer,取20u重新溶解的沉淀,进 行SDS-P A G E 电泳。
复性
1 ) 主要设备与试剂
2 mmol/L 还原型谷胱甘肽和0.2 mmol/L 氧化型谷胱甘肽。
2 ) 操作步骤
(1)每毫升脲-蛋白溶解液缓慢加入9m l 含 2mmol/L 还原型谷胱甘肽和0.2 mmol/L 氧化型谷胱甘肽的工作缓冲液。
⑵ 室 温 孵 育 2〜 4 h 。
融 合 蛋 白 的 纯 化 (以 G S T 标 签 融 合 蛋 白 的 纯 化 为 例)
1 ) 主要设备与试剂
摇床、离心机、超声波发生器、 L B 培养基、 IPTG(100 mmol/L )、 TritonX-100(10%)、5 0 % 的谷胱甘肽-琼脂糖珠、还原型谷胱甘肽洗脱液。
2 ) 操作步骤
(1)挑取一个菌落接种于 100 ml L B /A m p 培养基中,在 3 7 ℃ 摇 床 中 培 养 12〜 15 h。
⑵ 以 I : 1 0 的比例将此培养物稀释于 I L 新鲜的抗性L B 培养基中,分 入 2 个 2L的瓶中, 37℃下 培 养 lh。
(3)加lOOmmol/LIPTG 至终浓度 0.1 mmol/L ,继续培养 3〜 7 h 。
⑷ 室 温 、5OOOg离 心 IOmin收 集 细 胞 ,弃 上清。沉 淀 重 悬 于 10——20m l 冰 浴 的 PBS中。
(5) 将离心管置于冰浴中,用带直径5m m 探头的超声波发生器裂解细胞。
(6) 加 入 1 0 % 的 Triton X -100至浓 度 为 1 % 并混匀。室温、 10 OOOg离 心 5m i n 去除未裂解的细胞。
(7) 上 清 加 入 Iml 5 0 % 的谷胱甘肽-琼脂糖珠混悬液,室温下轻柔混勻2 min。加 50 ml冰 浴 的 P B S 洗涤液、混 勻 , 5 ( % 离 心 10s。重复 洗 涤 2 次 。用少量 (1〜 2 ml)冰 冷 的 PBS重悬琼脂糖珠,并 移 人一个1.5m l 微量离心管中。
(8) 室温、 5O O g 离 心 IOs收集琼脂糖珠,弃上清。加 Iml还原型谷胱甘肽洗脱融合蛋白质。轻 柔 混 勻 2 min, 500 g 离 心 10s,收集上清,重 复 洗 脱 2——3次 ,经 SDS-PAGE分析各部收集样品。洗脱的蛋白质分成小份,加 甘 油 至 终 浓 度 1 0 % ,储存于-70℃。同时测量吸光度确定融合蛋白的产量。
抗 体 的 制 备
一般来说,抗体制备主要分为两种!一种是用纯化的抗原免疫动物,获得多克隆抗体 ,这种方法在试验中比较常用;另一种是用杂交瘤技术制备单克隆抗体。
多 克 隆 抗 体 的 制 备
一般用纯化抗原免疫动物,可以得到特异性抗血清。免疫动物的抗原虽然进行了纯化 ,可每种抗原分子常有多个抗原决定簇,可以刺激动物产生多种质量不同的抗体,因此异种抗血清是由多种抗体组成的混合物,这类抗血清称为多克隆抗体,在实验研究中应用较广。我们以常见的蛋白质抗原免疫家兔制备抗血清为例进行介绍。
1 ) 所用设备与试剂
注射器、眼科剪、止血钳、引流管、无菌的玻璃瓶、家兔、弗氏完全佐剂、麻醉剂、叠氮钠(N a N 30.02%)。
2 ) 操作步骤
(1) 选择动物: 2.5〜 3k g 的健康家兔。
(2) 免疫动物:基础免疫所需抗原量为 0.5〜 lmg/kg,与弗氏完全佐剂以等体积乳化后做双后足掌注射及背部皮内多点注射。加强免疫,在第一次注射后1 〇天,用前次相同的抗原量与弗氏完全或不完全佐剂乳化,做腋窝淋巴结内注射,每 侧 a i ——〇.2m l ,剩余量做背部皮内多点注射, 1〜2周后,再重复注射一次可不加佐剂,抗原剂量适当增加,注射途径可多点皮下、肌肉或静脉内加强免疫。以 后 每 隔 1——2周 注 射 2——3次 ,末次注射后7——10 天试血,如果抗体效价不高,可再继续加强,直至达到要求的抗体效价(以双向扩散法检测,抗 体 效 价 在 1 : 3 2 以上),方可采血。
(3) 采 血 :颈动脉放血法。麻醉后将兔仰卧,四肢固定,充分暴露颈部,剪 毛 ,消毒 ,沿气管表面做皮肤直切口,暴露气管,在气管旁剥离动脉与神经束。用止血钳夹住动脉的近心端,用丝线结扎远心端,用眼科剪在两端被阻断的血管上剪一 「V 」形小口,向心插入引流管用丝线将插管与血管固定,放开近心端管钳,收集血液于清洁无菌的玻璃瓶中,置室温,待血块收缩完全后分离血清,加 最 终 浓 度 为 0.02% 的叠氮钠(N a N 3),小量分装,置-20℃冻存或冷冻干燥保存。
3 ) 注意事项
(1) 免疫用的动物多为哺乳类和禽类。哺乳类■主要有家兔、豚鼠、猪及猴等。禽类常用鸡。
(2) 免疫动物所用的抗原要求尽量纯化。
( 3 ) 目前用得最多的是弗氏佐剂,而弗氏佐剂又分为完全型和不完全型。
(4) 免疫动物需根据抗原的免疫原性强弱及抗原用量的多少,制定免疫方案。免疫的方案应包括注射的抗原量、途径、间隔时间和免疫次数。
单 克 隆 抗 体 的 制 备
单克隆抗体(monoclonal antibody, M c A b )有高度的均一性,特异性强。它是由抗原致敏的一个B 淋巴细胞增殖而来的细胞系所产生的抗体。这是因为单抗是来源于 — 个 B淋巴细胞的细胞株,而且其只对一种特定的抗原决定簇起反应,所以它具有高度均一性和特异性。不仅如此,杂交瘤细胞还能在体外或小鼠的腹腔内培养,产生大量抗体,易于标准化。
1 ) 所用设备与试剂
注射器、无菌培养皿、组织研磨器、计数器、恒温水浴、离心机、 2 4 孔板、二氧化碳孵箱、 B A L B /C 小鼠、弗氏完全佐剂、无菌生理盐水、 7 5 % 乙醇、 R P M I -1640培养液、5 % 小 牛 血 清 1640液 、台盼蓝、 5 0 % P E G 溶 液 、 2 0 % 小牛血清H A T 选择培养液。
2 ) 操作步驟
(1)动物免疫:常 选 用 B A L B /C 小 鼠 ,进行免疫,抗原用量根据所制备的抗血清的要求不同而异。常 用 12.5〜50 ug/100ul, 与等量弗氏完全佐剂混合,制成乳剂,用 200ul做动物背部皮内多点注射,如果注射量大,可做皮下注射 。 一 般间隔1——3周注射一次,共 3〜 5 次 ,末次接种后一周,切断小鼠尾巴尖部,放 血 ,检测抗血清的特异性和效价。选择产生高质量抗血清的小鼠作为脾细胞的来源。被选小鼠在试血后的 3〜6天,用同一剂量的抗原以无菌生理盐水溶解做静脉加强注射一次,使抗原刺激的B 淋巴细胞定位于脾脏。 3〜 4 天 后 ,处死动物,制备脾细胞。
⑵ 脾 细 胞 制 备 :将免疫的小鼠处死,浸 人 7 5 % 乙醇, 5m i n 换乙醇一次,在无菌条件下取出脾脏,用 R P M I -1640培养液冲洗表面血液,去包膜,将脾脏放入盛有培养液无菌培养皿中,剪 碎 移 入 15m l 的组织研磨器中,加 新 配 的 5 % 小 牛 血 清 1640液 ,挤压脾组织块,制备细胞悬液,收集细胞悬液静置片刻,待组织碎片下沉后,吸 取 上 含 细胞的悬液用5 % 小 牛 血 清 1640液 洗 2 次 ,沉淀再悬浮于Iml 培养液中并用台盼蓝染色,计 数 ,要求活细胞占9 0 % 以上。
(3)骨髓瘤细胞制备:从处于对数生长期的 N S -I 小鼠骨髓瘤细胞株培养液中取细胞2xl07〜 4xl07</sup 个 ,加 5 % 小牛血清1640液离心lOmin,去上清,将沉淀重悬计数。
⑷ 细 胞 融 合 :取 脾 细 胞 IO8和骨髓瘤细胞 2xl07777</sup个/m l , 饲养细胞可用小鼠腹腔巨噬细胞或未免疫小鼠脾细胞)混匀 ,分 配 到 2 4 孔 板 ,每孔加〇.5m l ,置 3 7 ℃ 含 5 % 〜 7 % 二氧化碳孵箱中培养。经 H A T 选择性培养后,融合的杂交瘤细胞生长良好, 而亲代的脾细胞和骨髓瘤细胞死亡。培 养 7〜 10天 后 ,在倒置显微镜下观察,见融合细胞已覆盖1/3〜 1/4孔底面积时,开始测抗体活性,应反复测定2〜 3 次 ,以确定抗体阳性孔。选出抗体阳性孔的细胞 , 一 部分做克隆化培养,一部分用液氮冷冻保存。
(5) 克隆化培养:常用有限稀释法 (limiting dilution)将抗体阳性孔的细胞进行准确计数 ,使细胞数达到10 个/m l ,然后根据所做9 6 孔板的数量,计 算 所 需 15% 小 牛 血 清 1640培养液与杂交瘤细胞的量,将细胞悬液分配到9 6 孔 板 ,每 孔 0.1 ml。使平均每孔有1个细 胞 ,再加入饲养细胞,一起 培 养 7〜 14天 ,待细胞克隆出现,并 覆 盖 1/3〜 1/4孔底时,再次检测培养液中的抗体活性。并将抗体阳性克隆化细胞再克隆培养 。 一 般至少应进行再克隆培养 2 次以上,按这种方法分配,将得到由单个细胞产生的克隆。
(6) 扩大培养:当找到抗体阳性克隆,应立即扩大培养,收 获 M c A b 。扩大培养制备 M c A b 的方法有:①抗体阳性细胞体外培养,收集培养液,但产量甚微,②腹水培养阳性细胞。具体操作是将无菌石蜡油0.5m l ,注入小鼠腹腔,造成无菌性^症 ,于 注 射 后 1〜 2 周 ,取 IO67</sup个抗体阳性细胞用〇.5m l 生理盐水稀释,注入小鼠腹腔,待产生大量腹水后,用注射器抽取腹水,每2〜3 天抽一次,可得到大量的 M c A b 。再经双向免疫扩散,免疫电泳及聚丙烯酰胺圆盘电泳,进行抗体鉴定,然后冷冻干燥,-20℃保存。
(7) 冷冻保存与复苏:冷冻应采用逐渐降温法,各实验室使用的方法大同小异。将细胞装入冰存管,加用小牛血清配制的1 〇%——15% 二甲亚砜(dimethyl sulfoxide, D M S O ),作为细胞保护剂。再将细胞从室温转到4℃ 、 -20℃ 、 -4〇℃ :,在-4O ℃ 放置6——2h处后直接放入液氮中。复苏时,将盛有细胞的冻存管从液氮中取出,立即投入 3 7 ℃水浴中,见已冻融迅速取出,将 细 胞 转 入 IOml离心管中,加 IOml 完全培养基, 1200r/m i n 离 心 5 min,然后将细胞转人培养瓶中培养。
3 ) 注意事项
(1) 作为免疫的小鼠应与骨髓瘤源自同一品系,这样克隆化的杂交瘤细胞,接种到小鼠才易于增殖,且容易获得高质量的单克隆抗体。
(2)挤压脾组织块,制备细胞悬液时只能挤压不能研磨。
(3) 用 来 制 备 M c A b 的 骨 髓 瘤 细 胞 是 B A L B /C 小 鼠 的浆细胞瘤,瘤细胞内缺乏>H G P R T 酶。 目前已筛选出许多不同的骨髓瘤细胞株。制 备 M cA b 应选择不分泌或不合成 小 鼠 Ig 重链、轻链的骨髓瘤细胞株,常用的有 NS-1、 SP2/0 和 653 等 。
(4) 检 测 抗 体 活 性 ,可用放射免疫法,免疫组织化学法、血凝法和细胞毒试验等手 段 。
细 胞 转 染
在研究中会需要将外源 D N A 导入不同类型的细胞,从而分析其表达并在细胞水平上研究它对细胞生长的影响。外 源 D N A 通常包括基因组 D N A 、克隆的目的基因和寡核苷酸等。实验中常用的方法主要分为两种:物理化学法和病毒载体法。在物化方法中除D E A E -葡聚糖介导法只适合瞬时转染外,其他各种物化方法既可用于瞬时也可用于稳定转 染 ;而不同的病毒载体法,有的适用于稳定转染 (如逆转录载体法),有的适用于瞬时转染 (如腺病毒载体法)。
实验中比较常用的是磷酸钙沉淀法和 DEAE-葡聚糖介导法、电穿孔法、脂质体介导法。
物 理 化 学 方 法
磷酸钙沉淀法
磷酸钙沉淀法的机制还不完全清楚,可 能 是 Ca2+ -D N A 结合物形成的颗粒沉积在培养细胞的表面,导致细胞的非特异性内吞作用。
H E P E S 缓冲溶液系统的磷酸钙沉淀法 一 一 用 该 方 法 转 染 的 D N A 既可以是基因组D N A 也可以是质粒 D N A 。
1 ) 所用设备与试剂
C O 2 培养箱、无菌培养皿、离心管、培养基、 CaCl2(2.5mol/L )、 2xHEP E S -NaCl2 溶液 (HBS 组 成 : 280 mmol/L N a C l , 10 mmol/L K C l , 1.5 mmol/L N a 2 H P 〇 4, 12 mmol/L 葡萄
糖 , 50 mmol/L H E P E S )
2 ) 操作步骤
⑴ 转 染 前 2 处 内 ,按 Ix lO 6 个细胞/培养皿的密度接种 IOOmm 的培养皿, 3 7 ℃、 5 %C O 2 培养过夜。在加入沉淀前 2〜 4 h ,换用新鲜的培养基培养细胞。
⑵ 将 需 要 转 染 的 D N A 用乙醇沉淀 (10——50ug/皿) , 空 气 中晾干。将 D N A 沉淀重悬于 450ul 灭菌水中,加 入 2.5m ol/L C aC l2 50ul。
(3) 取 2x H B S 500ul 于一无菌的锥形管中,一边 吹 打 2x H B S ,一边逐滴加入 D N A /CaCl2 溶液,随即祸旋 5s,将其在室温下静置 20 min 以形成沉淀。
(4) 将沉淀均勻加入长满细胞的 IOOm m 培养皿中,轻轻晃动。
(5) 在 37℃ 、 5 % C 〇 2 培养箱中培养细胞 4〜 16 h 。除去培养基, P B S 洗 细 胞 2 次 加IOml 完全培养基培养细胞。
(6) 对于瞬时转染,可在转染后 48——7211 收集细胞,并进行表达分析。对于稳定转染,让细胞生长 2 个倍増时间 (当被转染细胞是贴壁细胞时,倍增时间一般为 1 8 〜2 4 h ) 后分乂培养皿中进行培养。
3 ) 注意事项
⑴ 转 染 用 D N A 需进行纯化,不纯将影响转染效率。
(2) D N A -磷酸钙的比例不当会影响转染效率。
(3) 转染质粒 D N A 时可加入特定的基因组 D N A (载体 D N A ) 提高转染效率。
DEAE-葡聚糖介导法
二乙胺乙基葡萄糖简称 D E A E -葡聚糖,是在生理 p H 条件下带正电荷的多聚阳离子试 剂 ,它能促进靶细胞摄入外源 D N A ,其机制推测是 D N A 与 D E A E -葡聚糖结合易于被靶细胞内吞,并减少细胞内核酸酶对 D N A 的降解作用。该法只适用于瞬时转染,所需D N A 比磷酸钙沉淀法少,且不使用载体D N A 。
1 ) 主要设备与试剂
C O 2 培养箱; 无菌培养皿; 离心管;Tris 缓冲盐溶液 (T B S )(溶 液 A :NaCl 8 g ,KCl 0.38 g ,Na2 H P O 4 0.2 g , Tris 3 g ,用 H C l 调 节 p H 至 7.5, 定 容 至 100m l ,过滤除菌。溶 液 B :CaCl2 • 2 H 20 1.5 g , MgCl2 • 6 H 20 lg,定容至 100m l ,过滤除菌。取上述 IOmI A 液加入三 蒸 水 89m l ,再 加 Iml B 液体,即配制好 T B S ,使用前过滤除菌)。
2 ) 操作步骤
(1) 取 5xl05 个细胞接种于I O O m m 培养皿, 37℃、 5 % C 0 2 培 养 3 天 ,达 到 30%——50%融合。
(2) 乙醇沉淀 4 ug 质 粒 D N A ,空气干燥,溶 于 40 ul T B S 中。
(3) 吸去细胞培养上清液,用 I x PBS 洗涤细胞,加入完全培养基。
⑷ 边 摇 动 边 将 D N A 缓慢加入 80 ul预热至 37℃的 10 mg/ml D E A E -葡聚糖溶液中,再 将 120ulD N A /D E A E -葡聚糖混合液逐滴加人每个培养皿中,轻轻转动培养皿使其分布均 匀 ,置 3 7 ℃ 、 5 % C 0 2 培 养 4 h 。
(5) 吸 去 含 D N A /D E A E -葡聚糖的细胞培养基,加 人 5m l 含 1 0 % D M S O 的 lxPBS,室 温 静 置 Imin , 吸 去 含 D M S O 的 P B S , 用 5 ml P B S 漂洗细胞一次,加 IOml完全培养 基 继 续 培 养 48——72 h 。根据实 验 目 的 ,在适当的时间收获细胞,用于分析基因的瞬时 表 达 。
3 ) 注意事项
(1) D E A E -葡聚糖有较强的毒性,所以常采用低浓度 (1 〇〇 ——400 ug) 长时间 (4〜8h)的方案。
(2) 为了提高转染效率可以加入氯奎,它 可 以 保 护 D N A /D E A E -葡聚糖不被溶酶体降解。
电 穿孔法
该法是将细胞置于高压电场,在电脉冲的作用下,使细胞膜瞬间出现微小的孔洞,使 外 源 D N A 进入细胞。影响电穿孔法基因转移效率的因素有多个,其中电脉冲影响最大 。电穿孔转染的效率取决于受体细胞的类型、转染用的DNA构 象 等 。
1 ) 主要设备与试剂
低温离心机、电穿孔用的电击仪和电击池、含或不含有选择药物的完全培养基电穿孔缓冲液 [可根据实验所用的细胞类型,选用不同的电穿孔缓冲液,一般无血清和抗生素的细胞培养基,其电击条件为:电 容 960 uF ,电 压 250〜 450V (625——1125V /cm。
2 ) 操作步骤
(1) 培养靶细胞至对数生长中期, 4℃、 680 g 离 心 5m i n 收集细胞。
(2) 将细胞沉淀用预冷的电穿孔缓冲液重悬洗涤丨〜2 次 , 4 ℃ 、 6 8 % 离 心 5 min收集细胞。
⑶ 对 于 稳 定 转 染 ,将 细 胞 以 IxlO7 个/m l 的密度重悬于预冷的电穿孔缓冲液中。对于瞬时表达,可用更高密度的细胞。
⑷冰上放置所需数量的电击池,分 0.5m l 细胞悬液/池 。
(5) 将线状或超螺旋的 D N A (D N A 应经过纯化) 加入冰上放置的含细胞悬液的电击池中。握住电击池的两侧,晃动以混匀 D N A /细胞悬液,在冰上放置 5 min。
(6) 将电击池放人电击仪的池架上 (室温),用所设定的电压及电容值电击一次或多次 。电击的次数及电压和电容的设定随细胞类型而异,需进行摸索。
(7) 将电击池置于冰上 lOmin。
(8) 用非选择性完全培养基 2 0 倍稀释被转染细胞,并用该培养基洗电击池数次,以移出所有的被转染细胞。然 后 ,于 3 7 ℃ 、 5 % C O 2 培养细胞。
(9) 瞬时转染可于培养 48——72 h 后收获细胞,以分析基因的表达,稳定转染实验可采用含有特殊药物的选择性培养基进行抗性细胞克隆的筛选。
3 ) 注意事项
(I) D N A 浓度应控制在 1——40 ug/107 个细胞范围内,且 线 状 D N A 转染成功率较高。
⑵目前对于动物细胞进行电穿孔 ,一 般采用长脉冲/低电场方法。
脂 质 体 介 导 法
将需要转移的 D N A 等生物大分子包裹于脂质体,由于脂质体具有磷脂双层结构 (与细胞膜类似),因此可与细胞膜融合,将 D N A 等生物大分子转入耙细胞。单阳离子脂质 (如D O T M A 、 D O T A P 等) 及中性磷脂 (常见的为 D O P E ) 可 与 D N A 形成类脂复合物,从而使D N A 更有效地进人细胞。目前阳离子脂质转染试剂已商品化,转染率比其他转染方法更高 ,重复性更好。
1 ) 使用设备与试剂
培养皿、 C O 2 培养箱、培养基、商品化阳离子脂质转染试剂。
2 ) 操作步骤
(1) 以 1x 105 ——3x 105 个/孔 在 6 孔 板 或 35m m 培养皿中接种生长状况良好的靶细胞,于 37℃ 、 5 % C O 2 培养箱中培养 18〜 24 h ,至 细 胞 50%——80% 汇 片 。
(2) 在 E p 管中配制 D N A /脂质体复合物,体系如下:稀释 1——2 ug 质 粒 D N A 至 1(%1无 血 清 D M E M 培养基中。同 时 稀 释 2 —— 2 5ul阳 离 子 脂 质 于100ul 无 血 清 d m E M 培养基中,然后将二者混合,轻轻混匀。在室温下孵育 15〜 45 min,使 D N A 与阳离子脂质结合。
(3) 吸去细胞的原培养基,用 2m l 无血 清 D M E M 培养基洗细胞一次,在每一个孔或35m m 培养皿中加入 0.8m l 无 血 清 D M E M 培养基,然后逐滴加入上述 D N A /脂质体复合物 ,并使之均匀分布。于 3 7 ℃、 5 % C O 2 培养箱中培养 3〜 8 h ,一 般 为 5 h 。
(4) 每孔或直径 35m m 培养皿中加入 Iml 含 2 0 % 胎牛血清的 D M E M 完全培养基,置3 7℃ 、5 % C O 2 培养箱中再培养 18——24 h 。此过程中若有细胞毒性,必须更换新鲜的含 10%胎牛血清的 D M E M 完全培养基。
(5) 吸 去 DNA/脂质体复合物的 DMEM 完全培养基,每孔加人 2m l 新 鲜 的 含 10% 胎牛血清的 DMEM 完全培养基,再 培 养 24〜 48 h。
(6) 瞬时转染实验可在转染开始后 48——72 h 收获细胞,以分析基因的表达;稳定转染实验可于转染后 72 h ,将 细 胞 按 I : 1 0 分入含有特殊药物的选择性完全培养基进行抗性细胞克隆的筛选。
3 ) 注意事项
(1) 一般先加入无血清培养基,再加入未稀释的阳离子脂质体试剂。
(2) 如果待转染的细胞不耐受无血清培养基,可以先在无血清培养基中制备 D N A /脂质体复合物,然后将其加入含血清的完全培养基中进行转染。
病 毒 载 体 法
逆转录病毒载体法
基因组结构简单,易于操作,感染宿主范围广且效率高,但是只能感染分裂复制的细胞,滴度较低。此外,在包装细胞中,逆转录病毒载体有可能通过同源重组未野生型辅助病毒,有潜在的致癌危险性。
1 ) 使用设备与试剂
C O 2 培养箱、培养皿、培养板、克隆环、培养基、 H E P E S 缓冲盐溶液 (H B S ) 其组成如下 [137 mmol/L N a C l , 5 mmol/L K C l , 0.7 mmol/L N a 2 H P 〇 4, 6 mmol/L 葡萄糖, 21 mmol/L
H E P E S , 2mol/LCaCl2, 800 昭/m l (100x) 聚凝胺 (过滤除菌),含 15% 甘油的 H B S ]。
2 ) 操作步骤
⑴ 转 染 前 1 天 ,在 I O O m m 培 养 皿 中 1 : 5 或 I : 10 接种包装细胞培养至 10%〜 20%汇片。
⑵ 将 10 略含有药物抗性基因的逆病毒载体质粒 D N A 加 入 预 先 有 0.5 ml H B S 的试 管 中 ,再加入32 ul 2mol/LCaCl2,轻摇。室温下孵育 45min,直到形成细小的淡蓝色沉淀。
(3) 移去包装细胞的培养基,吸 取 H B S -D N A 沉淀轻加在培养皿中央。在超净工作台内让细胞接触 D N A 2Omin, 期间轻轻转动培养皿以均匀分散 D N A 溶液。加 IOmI 培养基 ,将细胞置于 3 7℃ 、 5 % C 0 2 培 养 4 h 。
(4) 吸尽培养基,轻 轻 加 入 2.5m l 室 温 的 H B S /甘油。将培养皿放回培养箱中孵育1.5〜 3.5 min。快速 弃 除 H B S /甘 油 ,用 IOmI 培养基轻轻洗 2 次 ,力卩 5m l 含有血清的培养基 ,培 养 18——24 h 移出培养基,用 0.45 um 的 滤 器 过 滤 ,获得含有短暂产生的病毒;将其保存于- 7 0 ℃或- 8 0 ℃。
(5) 在转染的细胞中加IOml培养基,培养 2〜3 天。
(6) 在 感 染 前 1 天 ,将包装细胞系 [与第⑴步所用的细胞系的 env 类 型 不 同 ] 按 I : 10至 1 : 2 0 传代。
(7) 移去所要感染的包装细胞系的培养基。加病毒 [从第 (5) 步来] 如下,对于一只I O O m m 的培养皿,稀 释 0.1——1.0m l 病毒液至终体积 3——5m l ,并 加 800ug/m l 的聚凝胺至对 于 60mm的 培养皿 ,稀 释 0.1〜 Iml 病毒液至终体积 2m l ,并 加 800ug/m l 的聚
凝胺至终浓度叫 8ug/ml。 3 7 ℃ 、 5 % C 0 2 培 养 I h 以上。
(8) 在 感 染 后 2——3 天 ,按 I : 10 或 1 : 2 0 将感染的细胞传代,加选择性培养基培养 3天 ,换用新鲜的选择培养基,再培养 4——7 天直至可见到细胞克隆。
(9) 用克隆环挑取分离良好的细胞克隆,每个克隆转移至 2 个 2 4 孔培养板或 6 孔培养板的培养孔中,生 长 至 5 0 % 〜 9 0 % 汇片。
(10) 更 换 1/2 体积的培养基,视包装细胞系不同而培养 1——3天,移出培养基,或对其进行滴度测定,或保存于-70℃ 或-80℃ 。
(11) 继续传代各产病毒细胞克隆,直至其可冻存或直至鉴别出最好的产病毒细胞。当鉴定出一个好的产病毒细胞克隆时,加 10%——15% DMSO 在液氮中冻存。
3 ) 注意事项
⑴ 如 果 感 染 持 续 时 间 超 过 6 h 甚至过夜,应使用更多体积的感染混合液,以防细胞脱水。
(2) 待感染靶细胞的接种数和感染混合液用量随培养皿的底面积而改变。
(3) 用几 种 量 的 病 毒 液 (如 0.1m l 、 0.4m l 、 1.0 ml) 感染几只培养皿,对于小鼠逆转录病毒和禽类逆转录病毒的 E 亚群需添加聚凝胺,对于禽类的其他亚群则不需要聚凝胺 。如果细胞对于聚凝胺不敏感 (大多数包装细胞都如此) 则延长培养时间,不会损害细 胞 。
⑷如果细胞对于聚凝胺不敏感,贝_ 培养基以增加体积。对 于 IOOmm 培养皿加培养基至终体积 10m l ,对于直径 60m m 培养皿加4 ml, 培 养 2〜 3 天。
腺病毒载体法
腺病毒可以感染分裂细胞和未分裂细胞,插 入 的 外 源 D N A 容量大。容易获得较高的滴度,病毒颗粒较稳定,但是其不足也比较明显,外源基因表达是暂时的,而且其体内应用容易引起免疫反应等。
1 ) 使用设备与试剂
C O 2 培养箱、培养板或培养皿、人胚肾 (HEK293) 细胞系、阳离子脂质体转染试剂。
2 ) 操作步骤
⑴ 在 6 孔培养板或直径 35m m 培养皿中接种约 5xl05 个 H E K 2 9 3 细胞, 5 % C O 2培养箱中培养24 h 。
(2) 提取重组病毒质粒 DNA 10|ag, 用 Pac I 酶切使之线性化。
⑶ 按 照 厂 家 说 明 用 lipofectamine(GIBCO 公司) 进行转染 293 细胞。
(4) 7〜 10天后用荧光显微镜进行观察,可以看到多数 293 细胞中有 G F P 的表达。
(5) 收取细胞,在- 7 0 ℃ / 3 7℃ 条件下反复冻融 4 次裂解细胞,离心取含病毒的上清再次 感 染 293 细胞以扩增重组病毒。
(6) 3 天后收集细胞,反复冻融 3 次。
(7) 取 400W 稀释液加至 293 细胞培养瓶中,于 3 7℃ 孵 育 3 h ,换新鲜培养基继续培养 18——24 h ,于荧光显微镜下计数 G F P 阳性细胞数,按以下公式计算病毒滴度 :
病毒滴度= G F P 细胞阳性数 X 病毒上清稀释倍数/〇.4 ml (pfu/ml)
3 ) 注意事项
收集毒液后可以进行纯化,用 CsCl 密度梯度超离心法进行病毒的纯化,将浓缩后的病毒储存液作不同比例的稀释。
定 位 研 究
定位研究的方法主要有荧光显微术、原位杂交、电子显微术。这里主要介绍通过荧光显微镜进行观察研究的方法,其试验策略是利用免疫学和非免疫学的荧光探针所具有的敏感性和特异性来揭示结构与功能的关系。
内源性 酶 检 测
β-半乳糖苷酶活性检测
炔半乳糖苷酶是经典的组织化学报道基因,其活性可以用多种底物来检测。最常用的 是 X -gal。当 々-gal 水 解 X -gal 分子的糖苷键时,会产生一种可溶性的、无色的吲哚单体 。随后,两个游离的吲哚基团无需酶促催化,即可自发氧化形成二聚体。最终形成的卤化靛青是一种极稳定且不溶于水的蓝色复合物。
1 ) 整块组织 (如胚胎) 的 X-gal 染色
⑴ 使 用 设 备 与 试 剂 :含 有 2m m 〇] y L MgCl2 的 P B S 、 2 % 〜 4 % 多聚甲醛、 X-gal 反应缓冲液 (35 mmol/L 亚铁氰化钾、 35 mmol/L 铁氰化钾、 2 mmol/L MgCl2、 0.02% N P -40、 0.01%
脱氧胆酸钠)、 X -gal 储存液 (40 mg/ml X -gal 溶 于 D M F )
(2) 操作步骤:
a. 将组织在冰上预冷的含有 2 mmol/L MgCl2 的 P B S 中剪碎。
b. 在 2%——4% 多 聚 甲 醛 中 冰 上 固 定 30m i n 到几个小时。
c. 用 P B S 漂洗几遍。
d. 用 X -gal 反应缓冲液稀释 X -gal 储存液,与组织在 3 7 1 孵 育 2——4 h 。
e.在 P B S 漂洗多次,直到漂洗液不再发黄为止。最好过夜漂洗。
f. 在明视野的显微镜下观察。
(3) 注意事项:
a. 固定时间不宜太久。
b. 在 X -gal 反 应 中 加 入 N B T 以替代铁离子,会形成紫色沉淀,这一反应比单用X -gal 反应更快更敏感。
2 ) 组织切片的 X-gal 染色
⑴ 使 用 设 备 与 试 剂 :明胶蔗糖包埋液 (7.5% 明胶、 15% 蔗糖、 0.05% 叠氮钠)、包被
载玻片的明胶溶液 (2 g 明胶, O.lg 硫酸铬钾溶于 200m l 水)、 4 % 多聚甲醛、 Gelvatol(①取


![2 ) 操作步骤 (1) 按上述方法进行(1)〜(7)操作。 (2) 与 1 : 5 0 稀释的碱性磷酸酶耦联的二抗室温孵育lh。 (3) 用 P B S 洗 3 次 ,每 次 5min。 ⑷ 在 室 温 用 bufferl洗 2 次 ,每 次 15min,随后在buffer2 中室温平衡Imin0 (5) 将细胞与2mlBCIP/N B T 溶液孵育,室温振荡显色。 (6) 定时用显微镜检查显色情况。通常的温育时间5〜 120min。 (7) 用 含 有 20mmol/L E D T A 的 P B S 冲洗以终止反应。将盖玻片封固进行显微镜 观 察 。 3 ) 注意事项 (1) 以上方法的优点在于酶/抗体复合物能够容易的渗透入细胞内并定位在其耙位, 有高度敏感性。 (2) 遇到的问题是内源性细胞酶活性的存在及对反应产物的检测结果模棱两可。 6 . 5 . 9 . 3 原 位 杂 交 原位杂交O hybridization)是以目标核酸的互补序列为探针,对细胞制品进行杂 交处理,使目标核酸可被呈现的技术。 6.5.9.3.1所用设备与试剂 杂交液、 4%多 聚 甲 醛 、 0.2111〇 1/1^11(:1、 0.1%〜0.3%1^〇 11乂-100、蛋白酶1^、乙醇、 5 0 % 甲酰胺、梓檬酸钠、 bufferl、 buffer 2、 buffer 3、 T E buffer。 6.5.9.3.2操作步骤 1 ) 探针的标记 选用地高辛(digoxingenin)标 记 的 d U T P , P C R 方法进行标记,玻璃奶方法回收目的 片段(地高辛标记的探针比非标记的P C R 产物稍大)。 2 ) 细胞片的处理 (1)取出培养约36h 的(HeLa)细胞爬片置于一个载玻片上, P B S (p H 7.2)洗 涤 3 次 ,空 气中放干。 ^ ⑵ 4 % 多聚甲醛室温固定10~30min。 (3) 0.2mol7L HCl 室温下作用 10min,0.1%~0.3% Triton X -100(PBS 配制)通透 15min。 ⑷ 用 含 0.2%甘氨酸的P B S 室 温 孵 育 15min。 ⑶ 蛋 白 酶 K [l| ig/m l , O.lmol/L Tris • Cl(p H 8.0)+50mmol/L E D T A (p H 8.0)配制]37X : 消 化 30min〇 (6) 含 0.2%甘氨酸的P B S 室 温 孵 育 lOmin。 (7) PBS-5mmol/L M g C l 2洗 2 次 ,各 lOmin。 (8) 4 % 多聚甲醛室温固定15min。 (9) PBS-5mmol/L MgCl2洗 2 次 ,各 15min。 (1〇 )逐级乙醇脱水(70%、 8 0 % 、 9 0 % 、 9 5 % 、 100%各 5min),空气中干燥。此玻片 可直接用于杂交,也可保存于4T 备用。](http://img.dxycdn.com/trademd/upload/userfiles/image/2016/07/A1469090019577ffpvs4fas7png_small.jpg)

蛋白质的结构/结构域。早期 的 M 13 方法和常规的P C R 方 法 ,由于操作繁琐,意外突变多等缺陷,使定点突变的实验受到很大限制,现在实验中常用的是 Stratagene 公司的 QuikChange Site-directed Mutagenesis Kit0
1 ) 操作步骤
⑴ 准 备 突 变 的 质 粒 从 常 规 中 经 纯 化 试 剂 盒 抽 提 质 粒 。
⑵ 设 计 一 对 互 补 的 、带有目标突变的引物,和模板质粒退火后用 PfuTurbo 聚合酶「循环延伸」。 PfuTurbo 聚合酶在非链置换反应条件下使引物线性延伸,产生环状带缺刻的突变产物。
(3) D/m I 酶切延伸产物,由于原来的模板质粒来源于常规大肠杆菌,是 经 d a m 甲基化修饰的,对 I 敏感而被切碎。
⑷带突变序列的质粒由于没有甲基化而不被切开形成具缺刻的双链 D N A ,将其转化 至 X L -2 Blue 超级感受态细胞中,即可得到突变质粒的克隆。
2 ) 注意事项
(1) 循环延伸是指聚合酶按照模板延伸引物 , 一 圈 后 回 到 引 物 5’端终止,再经过反复加热退火延伸的循环,这个反应区别于滚环扩增,不会形成多个串联拷贝。用的是低次数的循环延伸而非 P C R ,有助于减少无意错配。只需要一次酶切和转化。
(2) D/m I 识别序列为甲基化的 G A T C , G A T C 在几乎各种质粒中都会出现,而且不止一次。
磷 酸 化
蛋白质的磷酸化在细胞生长和分化的调控中扮演着重要的角色。靶蛋白的磷酸化程度由它们的蛋白质激酶和磷酸酶的相对活性决定。而且这两类酶的平衡受到许多因素影响 ,如生长因子、激素、细胞周期等。因此研究蛋白质磷酸化水平对于深入了解其所参与的信号转导过程是非常重要的。蛋白质磷酸化的定量测定常用 32P 来完成 (斯佩科特等2001)。
标记
1 ) 所用设备与试剂
细胞培养瓶、液闪计数器、刮 刷 、低磷酸培养基 (无放射性磷酸盐)、裂解缓冲液[20 mmol/L Tris(p H 7.5)]、 2 mmol/L E D T A 和 0.5% S D S 、 N P -40、 DNase I。
2 ) 操作步骤
(1) 用 60m m 或 I O O m m 培养瓶,在完全培养基中培养细胞至所希望的密度。
⑵用预热的低磷酸培养基 (无放射性磷酸盐) 冲洗 2 次 ,然后加人含 0.5〜ImCi32 P/ml(ICi = 3.7xl010 Bq) 的低磷酸培养基 (50——100umol/L 无机憐酸)。
(3) 培养结束后,回 收 32 P 标记培养基,用 IOml Aquasol(New England Nuclear) 稀 释 ,取一等份用液闪计数器测定 32P 活性。测定培养基中磷酸浓度,从这两个数据计算标记培养基中 32P 比活性。
⑷ 用 无 二 价 离 子 的 P B S 清 洗 32P 标记的细胞2 次。每 个 I O O m m 培 养 皿 加 Iml 预热的裂解缓冲液在沸水浴中裂解细胞。用一次性细胞刮刷将细胞裂解物刮到一边,然后吸到 1.5m l 离心管,热 水 浴 l 〇 min。
(5) 冷却至室温。
(6) 离心留上清,加 仰 - 4 0 至 2 % ,加 MgCl2至5mmol/L。加无蛋白酶的DNaseⅠ 与RNase 各 0.I m g ,室温放置 20 min
3 ) 注意事项
(1) 标准组织培养基有 l——2 mmol/L 磷酸盐。为 提 高 32P 标记效率,通常将培养基中磷酸盐浓度降至 5 〇 umol/L 或者更低。标记时间决定于胞内 A T P 库与培养 基 中 32 P 比活性达到平衡的快慢。
(2) 对于长时间培养,为避免类似成纤维细胞这样的快速分裂细胞生长减慢,培养基中无机磷酸浓度应保持在 50〜 100 mmol/L 。
(3) 测定培养基中磷酸浓度时也可用高效液相层析从其他 32P 标记的分子中分离A T P ,直接测胞内 [Y-32P] A T P 的比活性。
⑷ 2——4 h 的短脉冲标记可以用来研究瞬时的蛋白质磷酸化。然 而 ,在脉冲标记实验中 ,蛋 白 质 32 P 标记的暂时提高,可能是磷酸化的提高,也可能因为磷酸转换增强,而蛋白质稱联的磷酸的量并没有改变。分别用含有或不含刺激物的样品,用双向凝胶电泳/W estern 印迹比较未磷酸化蛋白质对磷酸化酸性同工型的比率,就可以区别以上两种可
能性。
检 测 32 P 标记的蛋白质
放射性磷酸 32 P 发出高能 P 粒子。因此,可 以 用 X 射线片放射自显影检测在单向或者双向凝胶上分辨出来的32 P 标记的蛋白质。
1 ) 所用设备与试剂
Whatman 3M M 滤纸、液闪计数器、胶片。
2 ) 操作步骤
(1) 单向或双向凝胶电泳分辨 32 P 标记的样品。
(2) 在 Whatman 3M M 滤纸上吸干凝胶。用放射性墨水在凝胶边缘标几个点。
(3) 在完全黑暗或安全光下,在 X 射线片曝光夹里,将胶片放在凝胶与增感屏之间。确保胶片与凝胶增感屏的闪烁体面紧密接触。让凝胶与胶片夹在两块增感屏之间。用两块塑料板夹紧曝光夹,于- 7 0℃ 曝光。
(4) 冲 洗 X 射线片,显影。
(5) 依放射性记号,在读片机里将考马斯亮蓝染色胶与放射自显影图排列好,标出靶蛋白,拍照。
(6) 切下含祀蛋白的胶条,用液闪计数器检测磷酸化蛋白的放射性。
3 ) 注意事项
放射自显影光密度分析也可以测定蛋白质带的比活性。但是,基于放射自显影对放射性进行精确的定量,需要合适的曝光时间,而且受到胶片狭窄的线性反应期的限制。如 果 有 P-扫描仪,可以用磷光成像板对32 P 放射性进行定量测定。
蛋 白 质 相 互 作 用
基因是相对静态的,而基因编码的产物—蛋白质,贝 U 是动态的。具有时空性和可

2 ) 实验步骤
(1) 将细胞裂解液与 5 0ul 的 5 0 % 谷胱甘肽琼脂糖球珠悬液和 25 飓 G S T 在 4℃ 颠倒混合 解 育 2 h 。
(2) 在微量离心机上以最大速度在 4 ¾ 离心混合物 2 min。
(3) 将上清转移到新的离心管中。
(4) 设定两个含等量上清及 5 0ul 谷胱甘肽琼脂糖球珠的微量离心管。在一管中加约10 ug 的 G S T 融合探针蛋白。两个反应中加入的探针和对照蛋白质的量应当是等摩尔的。将离心管在 4℃ 颠倒混合孵育 2 h。
(5) 在微量离心机上以最大速度离心样品 2 min。
(6) 在新的微量离心管中收集上清。这样样品可通过步骤 (10) 中 的 S D S -P A G E 进行分析。
(7) 用 Iml 冰冷的裂解液洗涤球珠。在微量离心机上以最大速度离心lmin。弃去上清 。重 复 洗 3 次 。
(8) (可选) 加人 50ul 20 mmol/L 的还原型谷胱甘肽 (在 50 mmol/L Tris • Cl p H 8.0 中) 到球珠中,洗 脱 G S T 融合蛋白及任何与其结合的蛋白质。在微量离心机上以最大速度离心2 min
(9) 将球珠 [来自步骤 (7)] 或洗脱蛋白质 [来自步骤 (8)] 与 等 体 积 S D S -P A G E 上样缓冲液混合。
(10) 将样品煮沸 4 min 并 作 S D S -P A G E 分析。
3 ) 结果判定
检 测 与 G S T 融合蛋白结合的蛋白质的方法取决于细胞裂解液是否被 35 S 标记以及试
验目的。如果目的是检测与融合蛋白结合的所有35 S标记的蛋白质,就在干胶仪上将胶抽 干 ,并 用 X 射线片曝光进行放射自显影。如果目的是检测特异性结合的蛋白质,就将蛋 白 质 从 S D S 聚丙烯酰胺凝胶转移至膜上,进行免疫印迹分析。如果目的是确定与融合蛋白结合的蛋白质的大小和丰度,而这些蛋白质是来自非放射性的裂解液,就用考马斯
亮蓝或硝酸银将胶染色。
免 疫 共 沉 淀
免疫共沉淀是以抗原和抗体之间的专一性作用为基础的研究蛋白质间相互作用的经典方法之一。在细胞的非变性条件下被裂解时,完整细胞内存在的许多蛋白质-蛋白质结合被保持下来。如果用蛋白质的抗体免疫沉淀该蛋白质,那么与该蛋白质在体内结合的另一蛋白质可能也沉淀下来。基于与蛋白质的生理性相互作用,蛋白质的免疫沉淀归类为免疫共沉淀。这种方法最常用于测定两种目标蛋白质是否在体内结合,但也可用于确定一种特定蛋白质的新的作用搭档。在保持蛋白质与蛋白质相互作用的条件下,收获和裂解细胞,从细胞提取液中特异性地免疫沉淀目的蛋白质,然后通过聚丙烯酰胺凝胶电泳分离免疫沉淀物。该方法的优点是相互作用的蛋白质都是经过翻译后修饰的,处于天然状态,蛋白质相互作用可以在自然状态下进行,能避免人为的影响,分离到天然状态的相互作用蛋白质 (Kay 2003)





水 和 大 约 3 0 粒无菌玻璃珠。② 把 5 个平板叠放在一起,紧握平板,猛烈地摇动平板直到菌落重新悬浮。③用无菌移液管从每个平板中吸取 5m l 酵 悬 液 。把细胞悬液集中在 50m l 的无菌锥形瓶中。④继续下面 5 个平板,重复①-③ I连 续 从 3 0 个平板中收获酵母细胞,结果得到总体积为 150m l 的酵母悬液,保 存 在 3 个 50m l 的瓶中。
ii.如果需要, 在每个装有酵母细胞的锥形瓶中加无菌 T E (p H 7.5 域无菌水至4〇 〜45m l ,振荡或翻转瓶中悬浮细胞。
iii. 使用台式离心机,室温、 1000——1500 g 离 心 5 min,弃去上清。
iv. 重 复 ii 和 iii。
v. 重新悬浮压紧的细胞沉淀于1 倍体积的无菌甘油溶液中,合并不同管的内容物 ,彻底混合。
Vi.分别转移 Iml 的细胞混合物于一系列无菌小离心管中, - 7 0℃ 冻存。
c.相互作用蛋白的筛选:
i. 解冻一份转化文库酵母细胞,用 C M (Gal-Raff)-Ura-His-Trp 培 养 基 按 1 : 10稀 释 , 3 0 ℃ 摇动培养酵母细胞 4 h 来诱导文库中 G A L l 启动子的转录。
ii.在适当数量的 100m m C M (Gal-Raff)-Ura-His-Trp-Leudropout 平板上分别培养 IO6 个细胞。
iii. 30℃ :培养平板 5 天。
iv.观察平板的生长,出现克隆时对其加以标记。
V. 第 5 天,涂布一^按^天出现的不同克隆分组的主平板 C M (Gal-Raff)-Ura-His_Tip。
vi. 3 0℃ 孵育平板直到菌落出现。
d. 阳性相互作用的确定:弟半乳糖苷酶活性和亮氨酸需求的检测。
e. 测定转录活化:①使用一个平头牙签分别在下面的 4 个平板上复制和主平板上相同的栅格。使用相同的牙签在 4 个平板上划线培养单克隆,尽量在含有 X-gal平板上划制粗的酵母线条,而在亮氨酸缺乏的平板上划细的酵母线条。②平板在 30℃:培 养 3〜 4 天 。
f.解释结果。如果具备下面的条件,菌落和它们包含的质粒被认为第一轮的阳性克 隆 :在 C M (Gal)-Ura-His-Trp 平板上的培养物 X -gal 呈现蓝色;在 C M (Glu)-U m -His-Trp 平板上的培养物 X -gal 分析呈现白色或仅是浅蓝色; 菌落在 C M (GalRaff)-Ura-His-Trp-Leu 平板上生长良好;菌落在 C M (Glu)-Ura-His-Trp-Leu 平板上生长缓慢或根本不能生长。
噬 菌 体 展 示 技 术
噬菌体展示技术是将表达多肽的基因与噬菌体表面蛋白编码的基因融合,以融合蛋白的形式表达在噬菌体表面的一种方法,最常用的表达系统是 M 1 3 噬菌体。将 c D N A 文库插入噬菌体载体进行表达后,所得到的总体为噬菌体展示库。为 了 得到与「诱 饵 」蛋白 相 作 用 的 「猎物」蛋白,可将展示库与固定化的「诱馆」蛋白相互作用,则 「猎 物 」蛋白被吸附下来,被吸附的重组噬菌体可经再感染而扩增,从而得到猎物蛋白的插入基
因 ,目前,噬菌体展示技术可用于鉴定和分析与其他蛋白质相互作用的蛋白质的表面特征 ;分离与特定氨基酸序列结合的新的配体;提高配体与靶结构之间相互作用的亲和力和特异性 (Brymora 2004)。
在微量滴定板上进行亲和筛选的方法,是将标靶蛋白固定在微量滴定板上,噬菌体重组肽库加到板上来选择相互作用的蛋白质。淘选过程可以再重复 2 次 ,以富集可结合的噬菌体减少非特异性结合噬菌体的回收。经 过 3 次的淘选,特异性连接的单克隆可通过 E L I S A 进行检测。连接上的可以在加入辣根过氧化物酶标记的抗噬菌体单克隆抗体及显色底物后呈现颜色。
噬菌体表面呈现库的构建主要包括以下的步骤:
(1) 首先对要呈现的目的基因片段进行随机酶切,并对酶切片段进行回收;
(2) 酶切片段和克隆接头进行连接,并插入噬菌体载体;
(3) 重组的噬菌体转人合适的宿主菌建库;
(4) 转化菌培养扩增后提取重组的噬菌体。
试剂与仪器
(I) 0.2mol/L 碳酸氢盐缓冲液 (Na2 H C O 3), p H 8.5;
⑵ 封 闭 缓 冲 液 :含 有 l%(m/V) BSA 的 PBS;
⑶ 洗 脱 缓 冲 液 :含 有 0.05%(W V )Tween-2 〇的 P B S ;
(4) 重组噬菌体展示库;
(5) 酸 洗 脱 缓 冲 液 : 50 mmol/L 甘氨酸-H C 1, p H 2.0;
( 6 ) 中和缓冲液: 0.2mol/LTris • Cl, pH7.5;
(7) 细菌: D H 5a F 培养过夜;
(8) 2x Y T 培养基以及顶层、底层琼脂:
胰蛋白膝 IOg
酵母膏 IOg
NaCl 5 g
溶 于 I L 的水中,高压灭菌。 4 ℃储 存 2 周 ;
(9) 底层琼脂:高压前加琼脂至终浓度为 1.5% (m/V);
(10) 顶层琼脂:高压前加琼脂至终浓度为0.8% (m /V);
(11) 阴性对照蛋白;
(12) 辣根过氧化物酶标记的抗噬菌体 M1 3 的单克隆抗体;
(13) 显色底物;
(14) 9 6 孔微量滴定板;
(15) 移液管;
(16) 5m l无菌管;
(17) 无菌牙签;
.
(18) 能读取微量滴定板的分光光度计;
(19) DNA 纯化测序设备。
方法
1 ) 微量滴定板包被
⑴ 纯 化 的 目 的 蛋 白 1〜 3 ug加 入 〇.5m l 的 0.2mol/L 的碳酸盐缓冲液 (p H 8.5 冲 混 合9 6 孔板中每孔加 1500,在室温下静置孵育 2 h ,少 数 蛋 白 质 可 以 代 过 夜 。
(2) 每孔加入 2 〇〇ul 的封闭缓冲液室温孵育 3 〇 min。
(3) 轻轻倒掉液体,在纸上轻叩两下,去除残留溶液。
(4) 每孔中加人 200叫洗脱缓冲液,洗 孔 3 次。
2) 选择相互作用的嗔菌体
⑴稀释重组噬菌体展示库,每 150ul 洗脱缓冲液含 1000 个噬菌体,之 后 取 150ul加入到孔中,室温孵育 2 h。
(2) 轻轻倒掉液体,在纸上叩干,去除残留溶液。
(3) 加 人 200|il 洗脱缓冲液,洗 孔 3 次。
3 ) 结合嗔菌体的回收
(1) 加 人 150ul 酸洗脱液,洗脱,在室温放置 lOmin,将 3 个孔的液体收集到一个干净 的 5m l 的塑料管中,每孔含有 1〇 3 ——1〇 8 个噬菌体颗粒。噬菌体也可以用〇.2m o l/ L 乙醇胺 (PH12.0) 洗 漆 ,然后中和。
(2) 加 人 200ul过夜培养的细菌,室温孵育 l〇 min。
(3) 加 人 3 ml 2x Y T 顶层琼脂培养基,混勻,倒 人 100个 15m m 的含有 2x Y T 底层琼脂培养基的培养皿中,在室温放置 5 min 凝 固 ,然后将板放于 3 7 ¾ 孵育过夜。(除了用板扩大噬菌体外,还可在液体培养基中扩大。将感染后的细菌的溶液加入到 5m l 的 2x Y T的肉汤中,转 移 至 50m l 的管中,于 37℃ 摇动过夜。)
⑷ 往 顶 层 琼 脂 上 加 人 5m l 洗脱缓冲液,用于洗提噬菌体颗粒,代 静 置 过 夜 。
(5) 取 5 0 0ul 噬菌体悬液13 200r/m i n 离 心 IOmin, 将上清移入新管中,这样就得到了噬菌体。加 400 ul 20%(m /V)P E G /2.5mol/LNaCl 到 1.6 ml 的上清中,冰浴 15 min,迅速
以 13 200r/m i n 离 心 5 min,再 用 2 0 0 0 洗脱缓冲液重悬。
( 6 ) 重复上述过程,以获得大量的噬菌体。每个循环收集到的噬菌体滴度为 IxlO11 pfu/ml
( 7 ) 收集到的噬菌体在 PB S 中 以 I : 100 稀释。在无菌的 5m l 的管中加入 200ul 过夜培 养 的 D H 5 〇 iF, 1〜 IOul 稀释后的噬菌体, 3m l 的 2x Y T 顶层琼脂培养,混勻。
(8) 将稀释的噬菌体接种在 2x Y T 底层琼脂培养基上, 3 7 ¾ 孵育过夜。
(9) 用无菌的牙签从噬菌斑挑取噬菌体,加 入 到 含 有 50ul 的过夜培养的 D H 5cxF 及lml2x Y T 培养基中。 37℃ 通风过夜培养生长。
4) ELISA 测定
(1) 9 6 孔板中一个孔用标靶蛋白包被,另一个用阴性对照蛋白包被,见 「 1) 微量滴定板包被」的步骤 (1)〜 (4)。 注意设置足够的孔用于检测「 3) 结合噬菌体的回收」中的步骤 (9) 得到的克隆。
⑵每对成对的孔中 (目标与对照伽入 1〇〇ul洗脱缓冲液和 2 5ul 的 「3) 结合噬菌体的回收」中的步骤(9) 得到的单克隆噬菌体的上清,室温孵育 2 h。
(3) 轻 轻 倒 掉 液 体 ,在纸上叩干,去除残留溶液。加 入 20〇 ul 洗脱缓冲液,洗孔3 次 。
⑷ 每 孔 加 l〇〇 ul 的抗噬菌体单克隆抗体(连有辣根过氧化物酶),以 1 : 5000 的比例稀 释 ,室 温 孵 育 lh。
(5) 轻轻倒 掉 液 体 ,在纸上叩干,去除残留溶液。加 入 200ul 洗 脱 缓 冲 液 ,洗孔3 次 。
(6) 每 孔 加 入 IOOul的显色底物,孵 育 3 〇 min。
(7) 4〇5n m 处测量光密度值。
5 ) 确 定 相 互 作 用 的 噬 菌 体 序 列
(1) 插入到重组噬菌体中的 D N A 纯化及测序。
(2) 将编码序列转化为肽序列。
噬菌体展示可用于:①鉴定和分析与其他蛋白质相互作用的蛋白质的表面特征;②分离与特定氨基酸序列结合的新配体;③提高配体与靶结构之间相互作用的亲和力和特异性。
蛋 白 质 结 构
蛋白质有成千上万种,不同的蛋白质具有不同的生物功能,生物功能的多样性是由蛋白质结构多样性所决定的。因此,蛋白质结构的研究占相当重要的地位。
蛋 白 质 一 级 结 构 的 测 定 方 法
蛋白质一级结构的测定有助于研究蛋白质的立体结构和蛋白质的功能,了解物种分子进化及鉴定分子病等。
1 ) 蛋 白 质 样 品 的 纯 化
蛋 白 质 样 品 与 等 体 积 S D S - P A G E 上 样 缓 冲 液 混 合 。将 样 品 煮 沸 4m i n 后进行SDS-PAGE 电泳。
2) 测 定 蛋 白 质 分 子 中 的 多 肽 链 的 数 目
通过测定蛋白质分子中 N 端 和 C 端残基的摩尔数及蛋白质的相对分子质量,可以确定蛋白质分子中肽链的数目,但应注意可能有封闭的末端,如乙酰化的 N 端和酰胺化 C端等。
3 ) 多肽链的分离
在测定一个蛋白质的结构以前,首先必须保证被测蛋白质的纯度,使结果准确可靠。其次要了解它的分子质量和亚基数,按照其亚基数将蛋白质分成几个多肽链。
(1) 肽链的拆开。蛋白质分子多肽链的连接有共价结合和非共价结合两种。要拆开以共价结合的一 S- S—连接的多肽链,必须采用的化学处理方法有:
a.过甲酸氧化:用氧化剂过甲酸断裂一 S—S— 。这个反应一般在〇丈下进行 2 h左 右 , 2 个 S 就全部能转变成磺酸基,这样被氧化的半胱氨酸称为磺基丙氨酸。
如果蛋白质分子中同时存在半胱胺酸,那么也会被氧化成磺基丙氨酸。此外甲硫氨酸和色氨酸也可被氧化,从而增加分析的复杂性。
b. 巯基乙醇还原:利用还原剂巯基乙醇亦可使蛋白质的一 S_s 一断裂。当高浓度的巯基乙醇在 P H 8.0 条件下室温保温几小时后,可以使一 s s 一定量还原。与此同时反应系统中还需要有 8mol/L 脲 或 6mol/L 盐酸胍使蛋白质变性,多肽链松散成为无规则的构型,此时还原剂就可作用于一 S—S— 。此反应是可逆的,
因此要使反应完全,巯基乙醇的浓度必须在 0.1——0.5moi/L。
c. Cldand 试剂的还原作用 Cleland 指出二硫赤藓糖醇 (dithioerythritol) 及二硫苏糖醇 (dithiothreitol)在氧化还原能力上是比较强的试剂,只 要 0.01mol /L 就能使蛋白质的一 S- S—还原,反应基本与巯基乙醇相似,且在许多球蛋白反应中,可以不用变性剂。 Cldand 试剂首先与蛋白质一 S- S—形成中间物,反应终了,还原剂被氧化形成一个稳定的六环化合物,蛋白质则被还原。蛋白质分子的几条肽链若以非共价键结合,则用尿素、盐酸胍等变性剂即可拆开。
(2) 蛋白质的多肽链被拆开后,将它分离纯化,一般多用凝胶过滤、离子交换、电泳等方法。
4 ) 测 定 多 肽 链 的 氨 基 酸 组 成
(1) 代 测 蛋 白 质 与 5.7 mmol/L 的盐酸在水解管中混合。并加入苯酚以防半胱氨酸和甲硫氨酸氧化,抽去空气,充满氮气, 110℃ 水 解 24 h ,或 在 150℃ 下快速水解 90 min。
(2) 水解得到的氨基酸与化学试剂反应,产生易被检测的氨基酸衍生物。氨基酸与茚三酮反应生成紫色化合物,在 570n m 处有吸收峰。脯氨酸和羟脯氨酸与茚三酮反应生成黄色化合物。
(3) 分离及检测。酸性条件下氨基酸带正电荷,是阳离子。在通过阳离子交换树脂时被吸附到柱上,再用洗脱液洗脱、收集。
5) N 端 和 C 端 氨 基 酸 残 基 的 测 定
N 端分析:①多肽的 a-氨基与异硫氰酸苯酯 (PITC) 试剂耦联;②在无水条件下用三氟乙酸酸裂解,形 成 A T Z -氨基酸;③ A T Z -氨基酸在三氟乙酸水溶液(25%) 中转化成稳定的 P T H -氨基酸;④通过薄层层析、气相色谱、高效液相谱及质谱进行氨基酸的鉴定。 C端分析: C 端自由羧基与乙酸酐反应生成混合酸酐,并进一步环化成曝唑酮, C 端® 唑酮在硫氰四丁胺的作用下,转化为乙内酰硫脲 (T H )。
(1) N 端测定:
a. 二硝基氟苯法(F D N B 、 D N F B ): D N P -氨基酸用有机溶剂抽提后,通过层析位置可鉴定它是何种氨基酸。 Sanger 用此方法测定了胰岛素的 N 端分别为甘氨酸及苯丙氨酸。
b. 氰酸盐法。
c. 二甲基氨基萘磺酰氯法。
(2) C 端分析:
a. 肼解法:这 是 测 定 C 端最常用的方法。将多肽溶于无水肼中, 1 0 0 ℃下进行反应 ,结果竣基末端氨基酸以游离氨基酸状释放,而其余肽链部分与肼生成氨基酸肼。这样羧基末端氨基酸可以采用抽提或离子交换层析的方法将其分出而进行分析。如果竣基末端氨基酸侧链是带有酰胺如天冬酰胺和谷氨酰胺,则肼解时不能产生游离的羧基末端氨基酸。此外肼解时注意避免任何少量的水解,以免释出的氨基酸混淆末端分析。
b. 羧肽酶水解法:羧肽酶可以专一性水解羧基末端氨基酸。根据酶解的专一性不同,可区分为羧肽酶 A 、 B 和 C 。应用羧肽酶测定末端时,需要事先进行酶的动力学实验,以便选择合适的酶浓度及反应时间,使释放出的氨基酸主要是 C端氨基酸。
6 ) 肽 段 的 氨 基 酸 序 列 分 析
E d m a n 降解的最大优越性是在水解除去末端标记的氨基酸残基时,不会破坏余下的多肽链。
⑴ 用 苯 异 硫 氰 酸 酯 (PITC)在 p H 9.0 的碱性条件下对蛋白质或多肽进行处理, PITC与肽链的N 端的氨基酸残基反应,形成苯氨基硫甲酰 (P T C ) 衍生物,即 F T C 肽 。
(2) P T C -肽用三氟乙酸处理, N 端氨基酸残基肽键被有选择地切断,释放出该氨基酸残基的噻唑啉酮苯胺衍生物。
⑶将该衍生物用有机溶剂(如氯丁烷) 从反应液中萃取出来,而去掉了一个 N 端氨基
酸残基的肽仍留在溶液中。萃取出来的噻唑啉酮苯胺衍生物不稳定,经酸作用,再进一步环化,形成一个稳定的苯乙内酰硫脲 (P T H ) 衍生物,即 P T H -氨基酸。留在溶液中的减少了一个氨基酸残基的肽再重复进行上述反应过程,整个测序过程现在都是通过测序仪自动进行。每一循环都获得一个 P T H -氨基酸,经 H P L C 可以鉴定出是哪一种氨基酸。当蛋白质中含有一个或多个半胱氨酸残基时,有时一对半胱氨酸残基会通过二硫键交联。在这种情况下测序,首先要经过处理 (如用过甲酸处理) 切断二硫键,然 后 再 进 行 Edman降解测。
7 ) 重 叠 法 确 定 肽 链 的 一 级 结 构
8 ) 蛋 白 质 分 子 中 二 硫 键 及 酰 胺 键 的 确 定 和 磷 酸
化 、糖 基 化 修 饰 点 的 定 位
(1) 蛋白质分子中二硫键的定位。
a. 蛋白质中由二硫键相连的肽链拆分:
i. 过甲酸处理:蛋白质中带游离巯基的半胱氨酸残基及由二硫键连接的半胱氨酸残基可被过甲酸氧化成黄基丙氨酸残基,将肽链拆分。
ii. 疏基乙醇或二巯基苏糖醇处理:用巯基乙醇或二巯基苏糖醇还原二硫键,再用碘乙酸烷基化,然后分别测定拆分得到的多肽链的一级结构。
b. 酶水解肽链未拆分的蛋白质,找出其中含有二硫键的肽段:蛋白质在微酸环境中进行 (P H 2.0〜 6.5)。 将肽段样品点样在正方形滤纸一角,第一方向碘用分离后,用过甲酸蒸气处理滤纸,然后滤纸旋转 90°,在与第一方向电泳的条件下进行第二方向电泳。由于两次电泳条件相同,不含二硫键的肽段在两次电泳中迁移率相同,最终聚集在正方形滤纸的对角线上。含有二硫键的肽段,在过甲酸处理后产生两个负电性增强的肽段,第二次电泳迁移率与第一次不同,不聚集在对角线上,据此可以加以区分。
(2) 酰胺基的定位。蛋白质的氨基酸组成分析可测定蛋白质中酰胺氮总量。
(3) 磷酸化修饰定位。用 ESI 质谱法或 MALDI 质谱法比较憐酸化前、后肽段的质谱可以定位磷酸化的位点。
(4) 糖基化位点的确定。肽段中糖计划修饰有两种类型,N-糖基化发生在门冬酰胺的侧链 N 原子上, O -糖基化发生在丝、苏氨酸侧链羟基的〇原子上,比较用糖苷酶水解前 、后糖肽的M A L D I M 谱便可以确定修饰位点 (鲁 子 贤 1982)。
蛋 白 质 二 级 结 构 的 测 定
1 ) 圆二色谱分析
圆二色谱分析是检测蛋白质二级结构信息的一种有用的方法。不同的二级结构对应与左旋光和右旋光的光吸收差值。通过分析圆二色谱曲线,可以估计二级结构螺旋、折叠以及无规则卷曲的比值。
样品的处理: Res0urseQ 和 R esourseS 离子交换柱分离的样品,低温中速离心浓缩至0.2 mg/ml,并将缓冲液换至测量缓冲液(浓度为 5 〇 m mol/LTris、 50 mmol/LNaCl、 5 mmol/LMgCl2、 p H 7.5 的缓冲液中含 0. 2 m g/mi) 中,直接用于测量。测 量 杯 长 度 为 0.2 cm , 溶液体 积 l 〇〇 ul。波长频率范围是 190〜 250nm。测量温度为 2 5 ℃
2 ) 动态光散射分析
动态光散射仪 (dynamic light scattering) 分 析 ,在 Protein Solution 公司的 Dyna Pro 仪器上运行。
样品处理: ResourseQ 和 ResourseS 离子交换柱分离的样品,低温中速离心浓缩至0.2 mg/ml,并将缓冲液换至测量缓冲液(浓 度 为 50 mm〇l/L Tris、 50 mmol/L NaCl、 5 mmol/LMgCl2、 pH7.5 的缓冲液中含〇.2 mg/ml 样品)中,取 2〇ul 蛋白质样品, 12 000r/min 高速
离 心 IOmin 后 ,用于测量。在 2 0 次的测量后,计算出溶液状态中的蛋白质的分子半径、分子质量以及主要的颗粒。测量温度为 25 丈 (Wei 1996)。
蛋 白 质 晶 体 空 间 结 构
X 射线衍射法分析蛋白质的晶体空间结构。
1 ) 材料与仪器
R I G U K U 转靶阳极 X 射线发生器安低温液氮控制系统、衍射数据收集的面探测仪、X 射线单色器。
2 ) 方法
X 射线晶体学研究通常采用的 X 射线波长与化学键键长相当,也与晶体内的原子间距离响应,约 为 O.lnm。一个晶体包括上亿个有序排列的基本单元。在晶胞中,原子核外电子组成的电子云对 X 射线产生衍射,由于晶体的周期性衍射束只在特定方向出现,电子云可以用傅里叶级数展开,其中每一项的大小与衍射强度相对应,可直接测量,而每一项的相位目前尚没有成熟的方法直接进行测量(牛 立 文 1998)。
(1) 结 晶 :需要通过大量的条件筛选和优化以恰好是蛋白质分子间的相互作用促成蛋白质分子形成高度有序的晶体,而不是随机聚合形成沉淀。这就要求溶液中的蛋白质处于合适的过饱和状态,并只形成少数的成核中心,使晶体能持续地慢慢生长成大单晶。
(2) 数据收集:通常利用 (单波长)X 射线光束照射在一定角度范围内旋转的蛋白质晶 体 ,同时记录晶体对 X 射线散射的强度。这些强度可转换为结构测定中的结构因子的振幅。
(3)相 角 的 测 定 :结构因子的振幅及相角不是直接相关的量。由于结构因子相角的全部信息在收集数据时丢失,因此必须通过其他途径来得到它们的数值。
(4)相角的改进 (优化):电子密度图的质量及其后的可解释性主要取决于相角的准确性 。有的情况下采用晶胞中不对称单元中的等同部分的电子密度平均,有可能大大地改善误差较大的起始相角。
(5) 电子密度图的解释:相位确定后,可开始计算电子密度图。若从电子密度图能跟踪出肽链走向和分辨出二级结构,则可能推出多肽链的三维折叠方式。进而根据氨基酸序列,就可能构建出原子坐标形式的蛋白质结构模型。
(6) 修 正 :考虑到以建立的立体化学资料的限制,根 据 X 衍射数据对初始的蛋白质分子模型进行修正 (Kensal 1998)。
增殖分化调控机制的研究
载 体 的 制 备
目的基因的克隆是分子生物学实验中的核心内容,起着承上启下的作用,通过克隆技术可实现目的基因的无限扩繁、长期保存的目的;利用该技术能实现基因-组、基因改造、构建工程菌和转基因动植物。基因克隆技术主要包括载体的获取、酶切、脱憐、连接和转化及筛选、鉴定等内容 (奥斯伯等 2001)。在干细胞增殖、分化调控机制的研究中常用到的克隆载体主要有以下两类:①宿主为大肠杆菌的细菌质粒;②动物细胞为宿主的动物病毒和以动物病毒为基础改造的穿梭质粒。
载体特点:①在宿主细胞中具独立复制能力;②带抗性等选择标记;③有合适的限制性内切核酸酶位点,可插人一定片段长度的外源 DNA 而不影响自身复制。
质 粒 提 取
质 粒 D N A 的提取方法比较常见,基本步骤多为三步:①细菌培养和质粒的扩增;②细菌菌体的裂解;③ 质 粒 DNA 的纯化。现在常用的是质粒提取试剂盒。该方法方快 捷 ,具体步骤可参考其使用手册。
注意:在 质 粒 DNA 提取中,质 粒 D N A 的质量和产量在很大程度上受培养条件的影响 ,通过一些实验发现,下列培养方式能获得较好的结果。①培养应用菌龄不超过 4 天的平板单菌落作为起始菌。②培养菌应在 37℃、 220r/min 下 生 长 16—— 18 h。③不能用经储藏的培养菌直接进行质粒提取制备。
D N A 片 段 的 酶 切
限制性内切核酸酶 (restriction endonuclease) 是一类能识别双链 DNA 分子中特异核苷酸序列的 DNA 水解酶,是体外剪切基因片段的重要工具。
1 ) 主要设备与试剂
离心机、恒温水浴、电泳仪、电泳槽、质粒、 TAE 缓冲液、限制性内切核酸酶
2 ) 操作步骤
用于质粒的酶切鉴定,一 般 是 20ul体积中含有0.2〜Iug D N A , 酶切反应体系为:
质 粒 DNA 10ul
缓冲液 (IOx) 2ul(双酶切时要选取两种酶都适合的缓冲液)
限制性内切核酸酶 1ul(双酶切时两种酶各加 1 ul)
双蒸水 7ul
⑴ 在 E p 管中加入 7 ul双蒸水。
⑵ 加 人 IOx 缓 冲 液 2ul 。
(3) 加 限 制 酶 1 ul。
⑷加入质粒 10 ul
(5) 离 心 ,混匀。
(6) 37℃水浴 l——2 h。
(7) 取 反 应 液 ,电泳鉴定。
3 ) 注意事项
⑴ 样 品 加 入 次 序 为 水 、缓冲液、 D N A ,最后为酶,不应颠倒。
(2) 加酶步骤要在冰浴中进行,在加酶前应先将水、缓冲液及待切 D N A 混匀。
(3) 反应体系的体积要尽量少 , 一 般 水 解 0.2〜 Iug D N A 时 ,应将体积控制在 20——30 ul内。保证限制酶活性不受抑制。
⑷为了控制反应体积和促进反应进行,要求模板 D N A 的浓度应很高,如 底 物 D N A浓度过低则应进行浓缩。
(5) 本实验中双酶切时,因两种酶的缓冲液盐浓度要求相同,所 以 ,可在同—反应体系中完成双酶切。但如进行双酶切时,两种酶所需缓冲液盐浓度要求不同,则不能将两种酶同时加入同一体系进行反应,此时可采取一种酶水解完后,进行有机溶剂抽提,沉淀、干燥后再复溶,然后进行第二种酶切。
D N A 片 段 回 收
回收是指从电泳介质中纯化出目的片段。在试验中待回收的片段主要有酶切片段和各 种 P C R 片段;回收的介质主要为琼脂糖;回收的方法主要有微量柱回收、玻璃奶回收。这里以玻璃奶法为例介绍 D N A 片段回收的方法。
1 ) 主要设备与试剂
离心机、烘箱、恒温水浴、玻璃奶、无水乙醇、溶胶液、漂洗液。
2 ) 操作步骤
(1) 从琼脂糖凝胶上切下目的条带放入 1.5 ml E p 管中,用 一 个 Tip 头捣碎。
⑵ 准 确 估 计 凝 胶 的 体 积 ,加 入 3 倍体积的溶胶液,室温放置 5m i n 或 50℃:保 温 3 min,其间轻摇 E p 管几次使胶完全溶化
。
⑶ 向 溶 胶 中 加 入 IOpl 玻璃奶,颠倒混勻,冰 浴 lOmin,并且间隔 2〜 3m i n 颠倒混匀一 次 , 12 000r/m i n 离 心 30s,吸弃上清。
(4) 将浓缩漂洗液按 3 : 7 与无水乙醇配成工作液,然后向玻璃奶中加入 2 5 0 ul ,用加样器吸漂洗液轻柔地将玻璃奶冲散混匀, 12 OOOr/m in 离 心 30s ,吸弃上清。
(5) 重复第 (4) 步一次。吸取完漂洗液后再离心 10s,用 Tip 头将最后一点儿漂洗液吸干 净 ,然后放置于 37℃ 温箱中干燥 20 min。
(6) 加 20ul 无菌水,用 Tip 头轻轻混勻, 60T ;水 浴 5 min, 12 OOOr/m i n 离 心 lmin,回收上清即为纯化的 P C R 片段。 -20℃ 储存。
3 ) 注意事项
(1) 玻璃奶使用之前一定要融化并充分混匀。
(2) 干燥后加无菌水后要充分吹打。
基 因 的 重 组 连 接
重组连接是指将目的基因用 D N A 连接酶连接在合适的质粒载体上,这是基因重组、基因改造的重要中间环节。连接反应可在溶液中进行也可在琼脂块中进行。在本部分仅以重组克隆为目的介绍相应的连接类型。在基因重组克隆中,外 源 D N A 与载体的连接是承上启下的一步操作,根据连接片段末端类型不同,连接方式有黏端连接、平端连接、平黏端连接及单碱基配对的 T 载体连接等。
主 要 设 备 与 试 剂
离心机、恒温设备、 T 4 D N A 连接酶、 10xT4 D N A 连接酶缓冲液。
操 作 步 骤
连接
1) 取干净、灭菌 E p 管,加入下列各试剂
T4 DNA 连接酶缓冲液 1ul
载 体 DNA 片段 1ng
插入目的片段 xng
连接酶 IU(平末端,如为黏端连接,则酶量可小于 1u)
加无菌水至 IOul
X = 目的片段与载体片段的分子比例 X 载 体 n g 数 X 目的片段的碱基数/载 体 n g 数
(目的片段/载体片段的分子比例一般为 3 : 1)
2) 连接反应
2 2 ℃ 保 温 3 h 或 4℃过 夜(Promega 连接酶,黏端连接);
或 22〜 2 6 ℃保 温 Ih (B R L 连接酶,黏端连接);
或 22℃保 温 4—— 16 h (Promega 连接酶,平端连接);
或 14℃ 保 温 16——24 h (B R L 连接酶)。
3 ) 注意事项
(1) 分子生物学实验中主要采用 T 4 D N A 连接酶。
(2) 在分子生物学实验中连接酶主要用于:①基因重组中载体与外源基因的连接;② 接 头 与 D N A 片段的连接,这种连接一般发生在文库构建、分子标记的 A F L P 及 差 ^表达的研究中等。
(3) 其中黏端连接的效率最高,黏端连接与平黏连接中外源 D N A 片段在载体中的插入有方向性。平端连接的效率最低,且载体发生自连的比例较髙,因此在克隆操作中为了克服平连的弊端,通常可通过:① T d T 酶在载体和外源 D N A 片段上转移一互补序列 (核苷酸投影法);②在载体和外源 D N A 片段上加上人工接头,变平连为黏连。
连接产物的转化
通过转化可以实现大量扩增质粒。转化的方法有多种,如电转化法、 CaCl2 法等。一般使用 CaCl2 法 ,从而改变细胞膜的结构,使 外 源 D N A 可以穿过细胞膜进入细胞,然后在选择性培养基中培养转化处理过的细菌,转化成功的细菌可以在抗性培养基上生长形成菌落。
1 ) 主要设备与试剂
摇床、离心机、恒温设备、 L B 培养液、 CaCl2(0.1mol/L )。
2 ) 操作步骤
(1) 从新鲜的平板上挑取一个 D H 5α 单克隆, 37℃摇振培养过夜。
(2) 按 I : 100 的比例将过夜培养的菌液加入新鲜的 L B 培养液中,于 37℃继续培养至 OD600为 0.3——0.4。
(3) 将菌液转移到预冷的 50m l 离心管中, 4 ℃ 、: IOOOg 离 心 15m i n 回收菌体。
(4) 倒出上清,将 管 倒 置 Imin 以使最后残留的痕量液体流尽。
(5) 用 IOml 冰预冷的 0.1mol/L CaCl2 重悬每份沉淀,同上离心收集菌体。
(6) 重复步骤⑷。
(7) 每 50m l 初始培养物用 2m l 冰预冷的 0.1mol/L CaCl2 重悬,此即感受态细菌。
(8) 取 200 叫感受态细菌和 D N A (≤10ul, ≤ 5 0 ng 伽入到一个冰冷的聚丙烯管中,轻轻旋转混匀,冰上放置 30 min。
(9) 将管放到 42 ℃水浴中,恰恰放置 90s,其间不要摇动。
(10) 快速将管转移到冰浴中,冷 却 l——2 min。
(11) 每 管 加 800ul L B 培养液, 37 ℃温和摇振培养 60m i n 使细菌复苏,并且表达质粒编码的抗生素抗性标记基因。
(12) 取 2 0 0 0 菌液涂在含 A m p 或 K a n 平板上,将平板置于室温直至液体被吸收,然 后 于 3 7 ℃ 倒置培养过夜。
3 ) 注意事项
(1) 热激温度和时间要严格控制好。
(2) 含 A m p 琼脂平板培养时间不应超过 16 h ,否则容易产生卫星菌落。
⑶ 如 果 是 与 T 载体的连接产物,则将菌液涂布在含 X -gal 和 I P T G 的 A m p 或 Kan平板上。
病毒载体的制备 (详见细胞转染部分)
1 ) 逆转录病毒载体的构建
⑴ 改 造 病 毒 基 因 组 ,切 除 野 生 型 的 娜 、 poZ、 e«v 结构基因,换为目的基因,保留包装信号和 L T R S 序列。
⑵将重组的病毒载体导入包装细胞,包装出有感染能力的逆转录病毒颗粒。
(3) 从包装细胞培养上清中获得包装好的缺陷型病毒,用它转导带有该病毒受体的祀细胞,使目的基因稳定整合至靶细胞染色体。
注意:①插入的目的基因不能超过 7kb , 否则逆转录病毒载体将不能有效包装。②外源基因导入靶细胞的效率很高,通 常 Im l 含病毒的培养基可感染 1 〇 6个觀细胞。
2 ) 腺病毒载体的构建 ( ;这部分已经商品化,不做详细介绍)
(1) 构建含外源目的基因的腺病毒穿梭载体。
(2) 制备重组质粒。
(3) 转 染 HEK293 细胞。
(4) 收获含重组腺病毒的上清进行滴度测定和纯化。
(5) 转染靶细胞。
注意: 一般插入外源 DNA 长度不得超过 2kb, 但是通过缺失部分病毒基因组可扩大外源 D N A 的插入容量。
蛋 白 质 的 制 备
在干细胞增殖、分化调控机制的研究中,常需要进行目的蛋白质的表达。首先我们通过载体制备将目的基因插入表达载体,然后再将该重组载体导入活细胞,以大量表达所需目的蛋白质。
外源基因可以在原核表达,也可以在真核细胞中进行表达。外源基因的原核表达是目的基因在原核细胞中以发酵的方式快速、高效的合成基因产物,常用于原核表达的细胞是大肠杆菌。其特点是:操作简便,而且在较短时间可以实现蛋白质的大量表达,因
此成为科研应用中蛋白质表达的主要宿主。但是大肠杆菌进行蛋白质表达也存在一些缺陷 :①真核蛋白在大肠杆菌中的表达没有得到正确的修饰;②在大肠杆菌中表达蛋白时常形成沉淀成为包含体。
诱导
常用表达用宿主菌株为 DE3 BL2 1 菌 ,先要准备合适的感受态细胞,取 质 粒 Ing 用无菌水稀释 5 0 倍 ,按前述转化步骤进行操作,筛选出阳性克隆后提质粒并经 D N A 序列测 定 ,确定无误后进行如下操作。
1 ) 主要设备与试剂
摇床、 SDS-PAGE 电泳仪、离心机、 LB 培养基、 IPTG、 Tris • HCI(20mmol/L pH8)、2xloading buffer
2 ) 操作步骤
⑴从新鲜的划线平板中挑取单克隆,接至一定体积的含合适抗生素的 L B 中, 37℃摇床振摇培养至OD600为 0.4〜 0.6。
(2) 如果表达载体的原核启动子为 PL 启动子,则 在 30〜 32℃ 培养数小时,使培养液的 OD600达 0.4〜 0.6, 迅速使温度升至 4 2 ℃ 继 续 培 养 3——5h。如果表达载体的原核启动子为 ta c 等 ,则 37T 培养细菌数小时达到对数生长期后加 IPTG至终浓度为lmmol/L,继续 培 养 3——5 h。
(3) 取 出 Im l菌液做分析用, 1 〇〇〇g发离心 lmin,沉 淀 加 l 〇〇ul 2xloadingbuffer, IOO℃加 热 3 min, 上 样 进 行 SDS-PAGE。
⑷ 将 摇 瓶 置 于 冰 上 5 min。4℃ 、 5 000g 离 心 5m in 收集菌体,重悬细胞于〇.2 5 倍体积预冷的2 〇 mol/L Tris HCl(pH 8.0) 中,离心。菌体保存于-7〇℃冻存以备分离基因表达产物所用。
3 ) 注意事项
不同的表达质粒,因启动子不同,诱导表达方法并不完全相同,可根据具体情况而定。
细 菌 的 裂 解
细胞破碎是提取胞内产物的关键性步骤。常用的研究方法有酶溶法和超声破碎法。
溶法
酶溶法就是用生物酶将细胞壁和细胞膜消化溶解的方法。常用的溶解酶有溶菌酶(Iysozyme)
1 ) 主要设备与试剂
离心机,裂解缓冲液 (lmmol/L EDTA, 100 mmol/L NaCl,50 mmol/L Tris • Cl,p H 8 . 0),蛋白酶抑制剂 (50 mmol/L ) , 溶菌酶(10m g /ml),脱氧胆酸, D N a SeI(lmg/ml)。
2 ) 操作步骤
(1) 4℃ 、 500 g 离 心 15 min,收集诱导表达的细菌(1000 ml)。
(2) 弃上清液,每克菌加3m l 裂解缓冲液,悬浮沉淀。
(3)每 克 菌 加 3ul 5Ommol/L 的 蛋 白 酶 抑 制 剂 苯 甲 磺 酰 氟 (phenylmethyIsulfonylfluoride, PMSF), 80ul 溶菌酶(10m g /ml),搅拌 20 min。
(4) 在搅拌下,每克菌加 4m g 脱氧胆酸。
(5) 溶 液 变 得 黏 稠 时 每 克 菌 加 20ul DNaseI(lmg/ml)。
(6)室温放置至溶液不再黏稠。
3 ) 注意事项
步骤 (2)〜 (4) 在冷室中进行。
超声破碎法
声 频 高 于 15〜 20kHz的超声波在高强度声能输入下可以进行细胞破碎,在处理少量样品时操作简便,液体量损失少,同时还可对染色体 D N A 进行剪切,大大地降低了液体的黏度,有利于后续目标产物的分离。
1 ) 主要设备与试剂
离心机、 SIDS-PAGE 电泳仪、 T E buffer、 2xloading buffer。
2 ) 操作步骤
⑴收集诱导表达的细菌(IOOOml),4℃ 、 500 g 离 心 15 min。
⑵ 弃 上 清 液 ,每克菌加3 ml T E buffer。
(3) 根据厂家提供的超声波仪的数据进行破菌。
⑷ 10 000g 离 心 15 min, 分别收集上清液和沉淀。
(5) 取 少 量 上 清 液 和 沉 淀 加 入 等 体 积 的 2xloading b u ffe r, 进 行 SDS-PAGE电泳。
3 ) 注意事项
(1) 超声破碎与声频、处理时间、细胞浓度、菌种类型等许多因素有关,实验时要根据具体情况掌握。
(2) 超声波破菌前,细胞经数次冻溶后更容易破碎。
包 含 体 的 分 离 、 溶 解 和 复 性
在以大肠杆菌为宿主菌高效表达外源基因时,表达蛋白常常在细胞质内聚集,形成包含体 (inclusion body)。包含体形成后,表达蛋白不具有物理活性,因此必须溶解包含体对表达蛋白进行复性。由于包含体是致密的,易于沉淀,所以能用脱氧胆酸盐洗涤得到纯度较高的制备物。包含体离心沉淀后,可用尿素等洗涤。为获得可溶性的活性蛋白,必须要将洗涤过的包含体溶解,然后进行重折叠,最后用离子交换或亲和层析的方法对重折叠后的蛋白质进行精制。
分离
1 ) 主要设备与试剂
离心机、 SDS-PAGE 电泳仪、尿素、 Tris . ClCaimolZL, p H 8.5)、 2xloading buffer。
2 ) 操作步骤
(1)4℃,12 OOOg离心细菌裂解液 15 min。
(2) 弃上清液,每 克 菌 加 Iml 无菌水使悬浮,分 别 取 1 0 0 ul 分 装 4 个 管 ,其余备用。
⑶ 离 心 同 步 骤 (1)。
(4) 弃上清液,用 lOOul O.lmolTLTris • Cl(p H 8.5),含不同浓度尿素(0.5mol/L 、lmol/L )的溶液悬浮沉淀。
(5)离 心 。
(6) 分别吸出并保留上清液,用 100ul H2O 重新悬浮沉淀。
(7) 分别取10 叫上清液和重新悬浮的沉淀,加 10ul 2xloading buffer进 行 SDS-PAGE电泳。
包含体的溶解
1 ) 主要设备与试剂
SDS-PAGE 电泳仪,裂解缓冲液 (含 0.1 m m 〇 l/L P M S F ,8mol/L 尿 素 ,10 mmol/L D T T ),K H 2P O 4(50 mmoiyL, p H 10.7), E D T A (lmmol/L , p H 8.0), NaCl(50 mmol/L )
2 ) 操作步骤
(1) 用 IOOul裂解缓冲液溶解包含体。
(2) 室 温 放 置 1h
(3) 加 9 倍体积的 P A G E 电泳缓冲液,室温放置30 min, 用 K O H 保 持 p H 为 10.7。
⑷ 用 H C l 调 至 p H 8.0,最少在室温放置30 min。
(5) 室温、 10 000g离心15min。
(6) 吸出上清液并保存,用 100 ul IxSDS-Ioading buffer重新溶解沉淀。
(7) 取 10 ul 上 清 液 ,加 10ul 2xloading buffer,取20u重新溶解的沉淀,进 行SDS-P A G E 电泳。
复性
1 ) 主要设备与试剂
2 mmol/L 还原型谷胱甘肽和0.2 mmol/L 氧化型谷胱甘肽。
2 ) 操作步骤
(1)每毫升脲-蛋白溶解液缓慢加入9m l 含 2mmol/L 还原型谷胱甘肽和0.2 mmol/L 氧化型谷胱甘肽的工作缓冲液。
⑵ 室 温 孵 育 2〜 4 h 。
融 合 蛋 白 的 纯 化 (以 G S T 标 签 融 合 蛋 白 的 纯 化 为 例)
1 ) 主要设备与试剂
摇床、离心机、超声波发生器、 L B 培养基、 IPTG(100 mmol/L )、 TritonX-100(10%)、5 0 % 的谷胱甘肽-琼脂糖珠、还原型谷胱甘肽洗脱液。
2 ) 操作步骤
(1)挑取一个菌落接种于 100 ml L B /A m p 培养基中,在 3 7 ℃ 摇 床 中 培 养 12〜 15 h。
⑵ 以 I : 1 0 的比例将此培养物稀释于 I L 新鲜的抗性L B 培养基中,分 入 2 个 2L的瓶中, 37℃下 培 养 lh。
(3)加lOOmmol/LIPTG 至终浓度 0.1 mmol/L ,继续培养 3〜 7 h 。
⑷ 室 温 、5OOOg离 心 IOmin收 集 细 胞 ,弃 上清。沉 淀 重 悬 于 10——20m l 冰 浴 的 PBS中。
(5) 将离心管置于冰浴中,用带直径5m m 探头的超声波发生器裂解细胞。
(6) 加 入 1 0 % 的 Triton X -100至浓 度 为 1 % 并混匀。室温、 10 OOOg离 心 5m i n 去除未裂解的细胞。
(7) 上 清 加 入 Iml 5 0 % 的谷胱甘肽-琼脂糖珠混悬液,室温下轻柔混勻2 min。加 50 ml冰 浴 的 P B S 洗涤液、混 勻 , 5 ( % 离 心 10s。重复 洗 涤 2 次 。用少量 (1〜 2 ml)冰 冷 的 PBS重悬琼脂糖珠,并 移 人一个1.5m l 微量离心管中。
(8) 室温、 5O O g 离 心 IOs收集琼脂糖珠,弃上清。加 Iml还原型谷胱甘肽洗脱融合蛋白质。轻 柔 混 勻 2 min, 500 g 离 心 10s,收集上清,重 复 洗 脱 2——3次 ,经 SDS-PAGE分析各部收集样品。洗脱的蛋白质分成小份,加 甘 油 至 终 浓 度 1 0 % ,储存于-70℃。同时测量吸光度确定融合蛋白的产量。
抗 体 的 制 备
一般来说,抗体制备主要分为两种!一种是用纯化的抗原免疫动物,获得多克隆抗体 ,这种方法在试验中比较常用;另一种是用杂交瘤技术制备单克隆抗体。
多 克 隆 抗 体 的 制 备
一般用纯化抗原免疫动物,可以得到特异性抗血清。免疫动物的抗原虽然进行了纯化 ,可每种抗原分子常有多个抗原决定簇,可以刺激动物产生多种质量不同的抗体,因此异种抗血清是由多种抗体组成的混合物,这类抗血清称为多克隆抗体,在实验研究中应用较广。我们以常见的蛋白质抗原免疫家兔制备抗血清为例进行介绍。
1 ) 所用设备与试剂
注射器、眼科剪、止血钳、引流管、无菌的玻璃瓶、家兔、弗氏完全佐剂、麻醉剂、叠氮钠(N a N 30.02%)。
2 ) 操作步骤
(1) 选择动物: 2.5〜 3k g 的健康家兔。
(2) 免疫动物:基础免疫所需抗原量为 0.5〜 lmg/kg,与弗氏完全佐剂以等体积乳化后做双后足掌注射及背部皮内多点注射。加强免疫,在第一次注射后1 〇天,用前次相同的抗原量与弗氏完全或不完全佐剂乳化,做腋窝淋巴结内注射,每 侧 a i ——〇.2m l ,剩余量做背部皮内多点注射, 1〜2周后,再重复注射一次可不加佐剂,抗原剂量适当增加,注射途径可多点皮下、肌肉或静脉内加强免疫。以 后 每 隔 1——2周 注 射 2——3次 ,末次注射后7——10 天试血,如果抗体效价不高,可再继续加强,直至达到要求的抗体效价(以双向扩散法检测,抗 体 效 价 在 1 : 3 2 以上),方可采血。
(3) 采 血 :颈动脉放血法。麻醉后将兔仰卧,四肢固定,充分暴露颈部,剪 毛 ,消毒 ,沿气管表面做皮肤直切口,暴露气管,在气管旁剥离动脉与神经束。用止血钳夹住动脉的近心端,用丝线结扎远心端,用眼科剪在两端被阻断的血管上剪一 「V 」形小口,向心插入引流管用丝线将插管与血管固定,放开近心端管钳,收集血液于清洁无菌的玻璃瓶中,置室温,待血块收缩完全后分离血清,加 最 终 浓 度 为 0.02% 的叠氮钠(N a N 3),小量分装,置-20℃冻存或冷冻干燥保存。
3 ) 注意事项
(1) 免疫用的动物多为哺乳类和禽类。哺乳类■主要有家兔、豚鼠、猪及猴等。禽类常用鸡。
(2) 免疫动物所用的抗原要求尽量纯化。
( 3 ) 目前用得最多的是弗氏佐剂,而弗氏佐剂又分为完全型和不完全型。
(4) 免疫动物需根据抗原的免疫原性强弱及抗原用量的多少,制定免疫方案。免疫的方案应包括注射的抗原量、途径、间隔时间和免疫次数。
单 克 隆 抗 体 的 制 备
单克隆抗体(monoclonal antibody, M c A b )有高度的均一性,特异性强。它是由抗原致敏的一个B 淋巴细胞增殖而来的细胞系所产生的抗体。这是因为单抗是来源于 — 个 B淋巴细胞的细胞株,而且其只对一种特定的抗原决定簇起反应,所以它具有高度均一性和特异性。不仅如此,杂交瘤细胞还能在体外或小鼠的腹腔内培养,产生大量抗体,易于标准化。
1 ) 所用设备与试剂
注射器、无菌培养皿、组织研磨器、计数器、恒温水浴、离心机、 2 4 孔板、二氧化碳孵箱、 B A L B /C 小鼠、弗氏完全佐剂、无菌生理盐水、 7 5 % 乙醇、 R P M I -1640培养液、5 % 小 牛 血 清 1640液 、台盼蓝、 5 0 % P E G 溶 液 、 2 0 % 小牛血清H A T 选择培养液。
2 ) 操作步驟
(1)动物免疫:常 选 用 B A L B /C 小 鼠 ,进行免疫,抗原用量根据所制备的抗血清的要求不同而异。常 用 12.5〜50 ug/100ul, 与等量弗氏完全佐剂混合,制成乳剂,用 200ul做动物背部皮内多点注射,如果注射量大,可做皮下注射 。 一 般间隔1——3周注射一次,共 3〜 5 次 ,末次接种后一周,切断小鼠尾巴尖部,放 血 ,检测抗血清的特异性和效价。选择产生高质量抗血清的小鼠作为脾细胞的来源。被选小鼠在试血后的 3〜6天,用同一剂量的抗原以无菌生理盐水溶解做静脉加强注射一次,使抗原刺激的B 淋巴细胞定位于脾脏。 3〜 4 天 后 ,处死动物,制备脾细胞。
⑵ 脾 细 胞 制 备 :将免疫的小鼠处死,浸 人 7 5 % 乙醇, 5m i n 换乙醇一次,在无菌条件下取出脾脏,用 R P M I -1640培养液冲洗表面血液,去包膜,将脾脏放入盛有培养液无菌培养皿中,剪 碎 移 入 15m l 的组织研磨器中,加 新 配 的 5 % 小 牛 血 清 1640液 ,挤压脾组织块,制备细胞悬液,收集细胞悬液静置片刻,待组织碎片下沉后,吸 取 上 含 细胞的悬液用5 % 小 牛 血 清 1640液 洗 2 次 ,沉淀再悬浮于Iml 培养液中并用台盼蓝染色,计 数 ,要求活细胞占9 0 % 以上。
(3)骨髓瘤细胞制备:从处于对数生长期的 N S -I 小鼠骨髓瘤细胞株培养液中取细胞2xl07〜 4xl07</sup 个 ,加 5 % 小牛血清1640液离心lOmin,去上清,将沉淀重悬计数。
⑷ 细 胞 融 合 :取 脾 细 胞 IO8和骨髓瘤细胞 2xl07777</sup个/m l , 饲养细胞可用小鼠腹腔巨噬细胞或未免疫小鼠脾细胞)混匀 ,分 配 到 2 4 孔 板 ,每孔加〇.5m l ,置 3 7 ℃ 含 5 % 〜 7 % 二氧化碳孵箱中培养。经 H A T 选择性培养后,融合的杂交瘤细胞生长良好, 而亲代的脾细胞和骨髓瘤细胞死亡。培 养 7〜 10天 后 ,在倒置显微镜下观察,见融合细胞已覆盖1/3〜 1/4孔底面积时,开始测抗体活性,应反复测定2〜 3 次 ,以确定抗体阳性孔。选出抗体阳性孔的细胞 , 一 部分做克隆化培养,一部分用液氮冷冻保存。
(5) 克隆化培养:常用有限稀释法 (limiting dilution)将抗体阳性孔的细胞进行准确计数 ,使细胞数达到10 个/m l ,然后根据所做9 6 孔板的数量,计 算 所 需 15% 小 牛 血 清 1640培养液与杂交瘤细胞的量,将细胞悬液分配到9 6 孔 板 ,每 孔 0.1 ml。使平均每孔有1个细 胞 ,再加入饲养细胞,一起 培 养 7〜 14天 ,待细胞克隆出现,并 覆 盖 1/3〜 1/4孔底时,再次检测培养液中的抗体活性。并将抗体阳性克隆化细胞再克隆培养 。 一 般至少应进行再克隆培养 2 次以上,按这种方法分配,将得到由单个细胞产生的克隆。
(6) 扩大培养:当找到抗体阳性克隆,应立即扩大培养,收 获 M c A b 。扩大培养制备 M c A b 的方法有:①抗体阳性细胞体外培养,收集培养液,但产量甚微,②腹水培养阳性细胞。具体操作是将无菌石蜡油0.5m l ,注入小鼠腹腔,造成无菌性^症 ,于 注 射 后 1〜 2 周 ,取 IO67</sup个抗体阳性细胞用〇.5m l 生理盐水稀释,注入小鼠腹腔,待产生大量腹水后,用注射器抽取腹水,每2〜3 天抽一次,可得到大量的 M c A b 。再经双向免疫扩散,免疫电泳及聚丙烯酰胺圆盘电泳,进行抗体鉴定,然后冷冻干燥,-20℃保存。
(7) 冷冻保存与复苏:冷冻应采用逐渐降温法,各实验室使用的方法大同小异。将细胞装入冰存管,加用小牛血清配制的1 〇%——15% 二甲亚砜(dimethyl sulfoxide, D M S O ),作为细胞保护剂。再将细胞从室温转到4℃ 、 -20℃ 、 -4〇℃ :,在-4O ℃ 放置6——2h处后直接放入液氮中。复苏时,将盛有细胞的冻存管从液氮中取出,立即投入 3 7 ℃水浴中,见已冻融迅速取出,将 细 胞 转 入 IOml离心管中,加 IOml 完全培养基, 1200r/m i n 离 心 5 min,然后将细胞转人培养瓶中培养。
3 ) 注意事项
(1) 作为免疫的小鼠应与骨髓瘤源自同一品系,这样克隆化的杂交瘤细胞,接种到小鼠才易于增殖,且容易获得高质量的单克隆抗体。
(2)挤压脾组织块,制备细胞悬液时只能挤压不能研磨。
(3) 用 来 制 备 M c A b 的 骨 髓 瘤 细 胞 是 B A L B /C 小 鼠 的浆细胞瘤,瘤细胞内缺乏>H G P R T 酶。 目前已筛选出许多不同的骨髓瘤细胞株。制 备 M cA b 应选择不分泌或不合成 小 鼠 Ig 重链、轻链的骨髓瘤细胞株,常用的有 NS-1、 SP2/0 和 653 等 。
(4) 检 测 抗 体 活 性 ,可用放射免疫法,免疫组织化学法、血凝法和细胞毒试验等手 段 。
细 胞 转 染
在研究中会需要将外源 D N A 导入不同类型的细胞,从而分析其表达并在细胞水平上研究它对细胞生长的影响。外 源 D N A 通常包括基因组 D N A 、克隆的目的基因和寡核苷酸等。实验中常用的方法主要分为两种:物理化学法和病毒载体法。在物化方法中除D E A E -葡聚糖介导法只适合瞬时转染外,其他各种物化方法既可用于瞬时也可用于稳定转 染 ;而不同的病毒载体法,有的适用于稳定转染 (如逆转录载体法),有的适用于瞬时转染 (如腺病毒载体法)。
实验中比较常用的是磷酸钙沉淀法和 DEAE-葡聚糖介导法、电穿孔法、脂质体介导法。
物 理 化 学 方 法
磷酸钙沉淀法
磷酸钙沉淀法的机制还不完全清楚,可 能 是 Ca2+ -D N A 结合物形成的颗粒沉积在培养细胞的表面,导致细胞的非特异性内吞作用。
H E P E S 缓冲溶液系统的磷酸钙沉淀法 一 一 用 该 方 法 转 染 的 D N A 既可以是基因组D N A 也可以是质粒 D N A 。
1 ) 所用设备与试剂
C O 2 培养箱、无菌培养皿、离心管、培养基、 CaCl2(2.5mol/L )、 2xHEP E S -NaCl2 溶液 (HBS 组 成 : 280 mmol/L N a C l , 10 mmol/L K C l , 1.5 mmol/L N a 2 H P 〇 4, 12 mmol/L 葡萄
糖 , 50 mmol/L H E P E S )
2 ) 操作步骤
⑴ 转 染 前 2 处 内 ,按 Ix lO 6 个细胞/培养皿的密度接种 IOOmm 的培养皿, 3 7 ℃、 5 %C O 2 培养过夜。在加入沉淀前 2〜 4 h ,换用新鲜的培养基培养细胞。
⑵ 将 需 要 转 染 的 D N A 用乙醇沉淀 (10——50ug/皿) , 空 气 中晾干。将 D N A 沉淀重悬于 450ul 灭菌水中,加 入 2.5m ol/L C aC l2 50ul。
(3) 取 2x H B S 500ul 于一无菌的锥形管中,一边 吹 打 2x H B S ,一边逐滴加入 D N A /CaCl2 溶液,随即祸旋 5s,将其在室温下静置 20 min 以形成沉淀。
(4) 将沉淀均勻加入长满细胞的 IOOm m 培养皿中,轻轻晃动。
(5) 在 37℃ 、 5 % C 〇 2 培养箱中培养细胞 4〜 16 h 。除去培养基, P B S 洗 细 胞 2 次 加IOml 完全培养基培养细胞。
(6) 对于瞬时转染,可在转染后 48——7211 收集细胞,并进行表达分析。对于稳定转染,让细胞生长 2 个倍増时间 (当被转染细胞是贴壁细胞时,倍增时间一般为 1 8 〜2 4 h ) 后分乂培养皿中进行培养。
3 ) 注意事项
⑴ 转 染 用 D N A 需进行纯化,不纯将影响转染效率。
(2) D N A -磷酸钙的比例不当会影响转染效率。
(3) 转染质粒 D N A 时可加入特定的基因组 D N A (载体 D N A ) 提高转染效率。
DEAE-葡聚糖介导法
二乙胺乙基葡萄糖简称 D E A E -葡聚糖,是在生理 p H 条件下带正电荷的多聚阳离子试 剂 ,它能促进靶细胞摄入外源 D N A ,其机制推测是 D N A 与 D E A E -葡聚糖结合易于被靶细胞内吞,并减少细胞内核酸酶对 D N A 的降解作用。该法只适用于瞬时转染,所需D N A 比磷酸钙沉淀法少,且不使用载体D N A 。
1 ) 主要设备与试剂
C O 2 培养箱; 无菌培养皿; 离心管;Tris 缓冲盐溶液 (T B S )(溶 液 A :NaCl 8 g ,KCl 0.38 g ,Na2 H P O 4 0.2 g , Tris 3 g ,用 H C l 调 节 p H 至 7.5, 定 容 至 100m l ,过滤除菌。溶 液 B :CaCl2 • 2 H 20 1.5 g , MgCl2 • 6 H 20 lg,定容至 100m l ,过滤除菌。取上述 IOmI A 液加入三 蒸 水 89m l ,再 加 Iml B 液体,即配制好 T B S ,使用前过滤除菌)。
2 ) 操作步骤
(1) 取 5xl05 个细胞接种于I O O m m 培养皿, 37℃、 5 % C 0 2 培 养 3 天 ,达 到 30%——50%融合。
(2) 乙醇沉淀 4 ug 质 粒 D N A ,空气干燥,溶 于 40 ul T B S 中。
(3) 吸去细胞培养上清液,用 I x PBS 洗涤细胞,加入完全培养基。
⑷ 边 摇 动 边 将 D N A 缓慢加入 80 ul预热至 37℃的 10 mg/ml D E A E -葡聚糖溶液中,再 将 120ulD N A /D E A E -葡聚糖混合液逐滴加人每个培养皿中,轻轻转动培养皿使其分布均 匀 ,置 3 7 ℃ 、 5 % C 0 2 培 养 4 h 。
(5) 吸 去 含 D N A /D E A E -葡聚糖的细胞培养基,加 人 5m l 含 1 0 % D M S O 的 lxPBS,室 温 静 置 Imin , 吸 去 含 D M S O 的 P B S , 用 5 ml P B S 漂洗细胞一次,加 IOml完全培养 基 继 续 培 养 48——72 h 。根据实 验 目 的 ,在适当的时间收获细胞,用于分析基因的瞬时 表 达 。
3 ) 注意事项
(1) D E A E -葡聚糖有较强的毒性,所以常采用低浓度 (1 〇〇 ——400 ug) 长时间 (4〜8h)的方案。
(2) 为了提高转染效率可以加入氯奎,它 可 以 保 护 D N A /D E A E -葡聚糖不被溶酶体降解。
电 穿孔法
该法是将细胞置于高压电场,在电脉冲的作用下,使细胞膜瞬间出现微小的孔洞,使 外 源 D N A 进入细胞。影响电穿孔法基因转移效率的因素有多个,其中电脉冲影响最大 。电穿孔转染的效率取决于受体细胞的类型、转染用的DNA构 象 等 。
1 ) 主要设备与试剂
低温离心机、电穿孔用的电击仪和电击池、含或不含有选择药物的完全培养基电穿孔缓冲液 [可根据实验所用的细胞类型,选用不同的电穿孔缓冲液,一般无血清和抗生素的细胞培养基,其电击条件为:电 容 960 uF ,电 压 250〜 450V (625——1125V /cm。
2 ) 操作步骤
(1) 培养靶细胞至对数生长中期, 4℃、 680 g 离 心 5m i n 收集细胞。
(2) 将细胞沉淀用预冷的电穿孔缓冲液重悬洗涤丨〜2 次 , 4 ℃ 、 6 8 % 离 心 5 min收集细胞。
⑶ 对 于 稳 定 转 染 ,将 细 胞 以 IxlO7 个/m l 的密度重悬于预冷的电穿孔缓冲液中。对于瞬时表达,可用更高密度的细胞。
⑷冰上放置所需数量的电击池,分 0.5m l 细胞悬液/池 。
(5) 将线状或超螺旋的 D N A (D N A 应经过纯化) 加入冰上放置的含细胞悬液的电击池中。握住电击池的两侧,晃动以混匀 D N A /细胞悬液,在冰上放置 5 min。
(6) 将电击池放人电击仪的池架上 (室温),用所设定的电压及电容值电击一次或多次 。电击的次数及电压和电容的设定随细胞类型而异,需进行摸索。
(7) 将电击池置于冰上 lOmin。
(8) 用非选择性完全培养基 2 0 倍稀释被转染细胞,并用该培养基洗电击池数次,以移出所有的被转染细胞。然 后 ,于 3 7 ℃ 、 5 % C O 2 培养细胞。
(9) 瞬时转染可于培养 48——72 h 后收获细胞,以分析基因的表达,稳定转染实验可采用含有特殊药物的选择性培养基进行抗性细胞克隆的筛选。
3 ) 注意事项
(I) D N A 浓度应控制在 1——40 ug/107 个细胞范围内,且 线 状 D N A 转染成功率较高。
⑵目前对于动物细胞进行电穿孔 ,一 般采用长脉冲/低电场方法。
脂 质 体 介 导 法
将需要转移的 D N A 等生物大分子包裹于脂质体,由于脂质体具有磷脂双层结构 (与细胞膜类似),因此可与细胞膜融合,将 D N A 等生物大分子转入耙细胞。单阳离子脂质 (如D O T M A 、 D O T A P 等) 及中性磷脂 (常见的为 D O P E ) 可 与 D N A 形成类脂复合物,从而使D N A 更有效地进人细胞。目前阳离子脂质转染试剂已商品化,转染率比其他转染方法更高 ,重复性更好。
1 ) 使用设备与试剂
培养皿、 C O 2 培养箱、培养基、商品化阳离子脂质转染试剂。
2 ) 操作步骤
(1) 以 1x 105 ——3x 105 个/孔 在 6 孔 板 或 35m m 培养皿中接种生长状况良好的靶细胞,于 37℃ 、 5 % C O 2 培养箱中培养 18〜 24 h ,至 细 胞 50%——80% 汇 片 。
(2) 在 E p 管中配制 D N A /脂质体复合物,体系如下:稀释 1——2 ug 质 粒 D N A 至 1(%1无 血 清 D M E M 培养基中。同 时 稀 释 2 —— 2 5ul阳 离 子 脂 质 于100ul 无 血 清 d m E M 培养基中,然后将二者混合,轻轻混匀。在室温下孵育 15〜 45 min,使 D N A 与阳离子脂质结合。
(3) 吸去细胞的原培养基,用 2m l 无血 清 D M E M 培养基洗细胞一次,在每一个孔或35m m 培养皿中加入 0.8m l 无 血 清 D M E M 培养基,然后逐滴加入上述 D N A /脂质体复合物 ,并使之均匀分布。于 3 7 ℃、 5 % C O 2 培养箱中培养 3〜 8 h ,一 般 为 5 h 。
(4) 每孔或直径 35m m 培养皿中加入 Iml 含 2 0 % 胎牛血清的 D M E M 完全培养基,置3 7℃ 、5 % C O 2 培养箱中再培养 18——24 h 。此过程中若有细胞毒性,必须更换新鲜的含 10%胎牛血清的 D M E M 完全培养基。
(5) 吸 去 DNA/脂质体复合物的 DMEM 完全培养基,每孔加人 2m l 新 鲜 的 含 10% 胎牛血清的 DMEM 完全培养基,再 培 养 24〜 48 h。
(6) 瞬时转染实验可在转染开始后 48——72 h 收获细胞,以分析基因的表达;稳定转染实验可于转染后 72 h ,将 细 胞 按 I : 1 0 分入含有特殊药物的选择性完全培养基进行抗性细胞克隆的筛选。
3 ) 注意事项
(1) 一般先加入无血清培养基,再加入未稀释的阳离子脂质体试剂。
(2) 如果待转染的细胞不耐受无血清培养基,可以先在无血清培养基中制备 D N A /脂质体复合物,然后将其加入含血清的完全培养基中进行转染。
病 毒 载 体 法
逆转录病毒载体法
基因组结构简单,易于操作,感染宿主范围广且效率高,但是只能感染分裂复制的细胞,滴度较低。此外,在包装细胞中,逆转录病毒载体有可能通过同源重组未野生型辅助病毒,有潜在的致癌危险性。
1 ) 使用设备与试剂
C O 2 培养箱、培养皿、培养板、克隆环、培养基、 H E P E S 缓冲盐溶液 (H B S ) 其组成如下 [137 mmol/L N a C l , 5 mmol/L K C l , 0.7 mmol/L N a 2 H P 〇 4, 6 mmol/L 葡萄糖, 21 mmol/L
H E P E S , 2mol/LCaCl2, 800 昭/m l (100x) 聚凝胺 (过滤除菌),含 15% 甘油的 H B S ]。
2 ) 操作步骤
⑴ 转 染 前 1 天 ,在 I O O m m 培 养 皿 中 1 : 5 或 I : 10 接种包装细胞培养至 10%〜 20%汇片。
⑵ 将 10 略含有药物抗性基因的逆病毒载体质粒 D N A 加 入 预 先 有 0.5 ml H B S 的试 管 中 ,再加入32 ul 2mol/LCaCl2,轻摇。室温下孵育 45min,直到形成细小的淡蓝色沉淀。
(3) 移去包装细胞的培养基,吸 取 H B S -D N A 沉淀轻加在培养皿中央。在超净工作台内让细胞接触 D N A 2Omin, 期间轻轻转动培养皿以均匀分散 D N A 溶液。加 IOmI 培养基 ,将细胞置于 3 7℃ 、 5 % C 0 2 培 养 4 h 。
(4) 吸尽培养基,轻 轻 加 入 2.5m l 室 温 的 H B S /甘油。将培养皿放回培养箱中孵育1.5〜 3.5 min。快速 弃 除 H B S /甘 油 ,用 IOmI 培养基轻轻洗 2 次 ,力卩 5m l 含有血清的培养基 ,培 养 18——24 h 移出培养基,用 0.45 um 的 滤 器 过 滤 ,获得含有短暂产生的病毒;将其保存于- 7 0 ℃或- 8 0 ℃。
(5) 在转染的细胞中加IOml培养基,培养 2〜3 天。
(6) 在 感 染 前 1 天 ,将包装细胞系 [与第⑴步所用的细胞系的 env 类 型 不 同 ] 按 I : 10至 1 : 2 0 传代。
(7) 移去所要感染的包装细胞系的培养基。加病毒 [从第 (5) 步来] 如下,对于一只I O O m m 的培养皿,稀 释 0.1——1.0m l 病毒液至终体积 3——5m l ,并 加 800ug/m l 的聚凝胺至对 于 60mm的 培养皿 ,稀 释 0.1〜 Iml 病毒液至终体积 2m l ,并 加 800ug/m l 的聚
凝胺至终浓度叫 8ug/ml。 3 7 ℃ 、 5 % C 0 2 培 养 I h 以上。
(8) 在 感 染 后 2——3 天 ,按 I : 10 或 1 : 2 0 将感染的细胞传代,加选择性培养基培养 3天 ,换用新鲜的选择培养基,再培养 4——7 天直至可见到细胞克隆。
(9) 用克隆环挑取分离良好的细胞克隆,每个克隆转移至 2 个 2 4 孔培养板或 6 孔培养板的培养孔中,生 长 至 5 0 % 〜 9 0 % 汇片。
(10) 更 换 1/2 体积的培养基,视包装细胞系不同而培养 1——3天,移出培养基,或对其进行滴度测定,或保存于-70℃ 或-80℃ 。
(11) 继续传代各产病毒细胞克隆,直至其可冻存或直至鉴别出最好的产病毒细胞。当鉴定出一个好的产病毒细胞克隆时,加 10%——15% DMSO 在液氮中冻存。
3 ) 注意事项
⑴ 如 果 感 染 持 续 时 间 超 过 6 h 甚至过夜,应使用更多体积的感染混合液,以防细胞脱水。
(2) 待感染靶细胞的接种数和感染混合液用量随培养皿的底面积而改变。
(3) 用几 种 量 的 病 毒 液 (如 0.1m l 、 0.4m l 、 1.0 ml) 感染几只培养皿,对于小鼠逆转录病毒和禽类逆转录病毒的 E 亚群需添加聚凝胺,对于禽类的其他亚群则不需要聚凝胺 。如果细胞对于聚凝胺不敏感 (大多数包装细胞都如此) 则延长培养时间,不会损害细 胞 。
⑷如果细胞对于聚凝胺不敏感,贝_ 培养基以增加体积。对 于 IOOmm 培养皿加培养基至终体积 10m l ,对于直径 60m m 培养皿加4 ml, 培 养 2〜 3 天。
腺病毒载体法
腺病毒可以感染分裂细胞和未分裂细胞,插 入 的 外 源 D N A 容量大。容易获得较高的滴度,病毒颗粒较稳定,但是其不足也比较明显,外源基因表达是暂时的,而且其体内应用容易引起免疫反应等。
1 ) 使用设备与试剂
C O 2 培养箱、培养板或培养皿、人胚肾 (HEK293) 细胞系、阳离子脂质体转染试剂。
2 ) 操作步骤
⑴ 在 6 孔培养板或直径 35m m 培养皿中接种约 5xl05 个 H E K 2 9 3 细胞, 5 % C O 2培养箱中培养24 h 。
(2) 提取重组病毒质粒 DNA 10|ag, 用 Pac I 酶切使之线性化。
⑶ 按 照 厂 家 说 明 用 lipofectamine(GIBCO 公司) 进行转染 293 细胞。
(4) 7〜 10天后用荧光显微镜进行观察,可以看到多数 293 细胞中有 G F P 的表达。
(5) 收取细胞,在- 7 0 ℃ / 3 7℃ 条件下反复冻融 4 次裂解细胞,离心取含病毒的上清再次 感 染 293 细胞以扩增重组病毒。
(6) 3 天后收集细胞,反复冻融 3 次。
(7) 取 400W 稀释液加至 293 细胞培养瓶中,于 3 7℃ 孵 育 3 h ,换新鲜培养基继续培养 18——24 h ,于荧光显微镜下计数 G F P 阳性细胞数,按以下公式计算病毒滴度 :
病毒滴度= G F P 细胞阳性数 X 病毒上清稀释倍数/〇.4 ml (pfu/ml)
3 ) 注意事项
收集毒液后可以进行纯化,用 CsCl 密度梯度超离心法进行病毒的纯化,将浓缩后的病毒储存液作不同比例的稀释。
定 位 研 究
定位研究的方法主要有荧光显微术、原位杂交、电子显微术。这里主要介绍通过荧光显微镜进行观察研究的方法,其试验策略是利用免疫学和非免疫学的荧光探针所具有的敏感性和特异性来揭示结构与功能的关系。
内源性 酶 检 测
β-半乳糖苷酶活性检测
炔半乳糖苷酶是经典的组织化学报道基因,其活性可以用多种底物来检测。最常用的 是 X -gal。当 々-gal 水 解 X -gal 分子的糖苷键时,会产生一种可溶性的、无色的吲哚单体 。随后,两个游离的吲哚基团无需酶促催化,即可自发氧化形成二聚体。最终形成的卤化靛青是一种极稳定且不溶于水的蓝色复合物。
1 ) 整块组织 (如胚胎) 的 X-gal 染色
⑴ 使 用 设 备 与 试 剂 :含 有 2m m 〇] y L MgCl2 的 P B S 、 2 % 〜 4 % 多聚甲醛、 X-gal 反应缓冲液 (35 mmol/L 亚铁氰化钾、 35 mmol/L 铁氰化钾、 2 mmol/L MgCl2、 0.02% N P -40、 0.01%
脱氧胆酸钠)、 X -gal 储存液 (40 mg/ml X -gal 溶 于 D M F )
(2) 操作步骤:
a. 将组织在冰上预冷的含有 2 mmol/L MgCl2 的 P B S 中剪碎。
b. 在 2%——4% 多 聚 甲 醛 中 冰 上 固 定 30m i n 到几个小时。
c. 用 P B S 漂洗几遍。
d. 用 X -gal 反应缓冲液稀释 X -gal 储存液,与组织在 3 7 1 孵 育 2——4 h 。
e.在 P B S 漂洗多次,直到漂洗液不再发黄为止。最好过夜漂洗。
f. 在明视野的显微镜下观察。
(3) 注意事项:
a. 固定时间不宜太久。
b. 在 X -gal 反 应 中 加 入 N B T 以替代铁离子,会形成紫色沉淀,这一反应比单用X -gal 反应更快更敏感。
2 ) 组织切片的 X-gal 染色
⑴ 使 用 设 备 与 试 剂 :明胶蔗糖包埋液 (7.5% 明胶、 15% 蔗糖、 0.05% 叠氮钠)、包被
载玻片的明胶溶液 (2 g 明胶, O.lg 硫酸铬钾溶于 200m l 水)、 4 % 多聚甲醛、 Gelvatol(①取
208,209-212
蛋白质的结构/结构域。早期 的 M 13 方法和常规的P C R 方 法 ,由于操作繁琐,意外突变多等缺陷,使定点突变的实验受到很大限制,现在实验中常用的是 Stratagene 公司的 QuikChange Site-directed Mutagenesis Kit0
1 ) 操作步骤
⑴ 准 备 突 变 的 质 粒 从 常 规 中 经 纯 化 试 剂 盒 抽 提 质 粒 。
⑵ 设 计 一 对 互 补 的 、带有目标突变的引物,和模板质粒退火后用 PfuTurbo 聚合酶「循环延伸」。 PfuTurbo 聚合酶在非链置换反应条件下使引物线性延伸,产生环状带缺刻的突变产物。
(3) D/m I 酶切延伸产物,由于原来的模板质粒来源于常规大肠杆菌,是 经 d a m 甲基化修饰的,对 I 敏感而被切碎。
⑷带突变序列的质粒由于没有甲基化而不被切开形成具缺刻的双链 D N A ,将其转化 至 X L -2 Blue 超级感受态细胞中,即可得到突变质粒的克隆。
2 ) 注意事项
(1) 循环延伸是指聚合酶按照模板延伸引物 , 一 圈 后 回 到 引 物 5’端终止,再经过反复加热退火延伸的循环,这个反应区别于滚环扩增,不会形成多个串联拷贝。用的是低次数的循环延伸而非 P C R ,有助于减少无意错配。只需要一次酶切和转化。
(2) D/m I 识别序列为甲基化的 G A T C , G A T C 在几乎各种质粒中都会出现,而且不止一次。
磷 酸 化
蛋白质的磷酸化在细胞生长和分化的调控中扮演着重要的角色。靶蛋白的磷酸化程度由它们的蛋白质激酶和磷酸酶的相对活性决定。而且这两类酶的平衡受到许多因素影响 ,如生长因子、激素、细胞周期等。因此研究蛋白质磷酸化水平对于深入了解其所参与的信号转导过程是非常重要的。蛋白质磷酸化的定量测定常用 32P 来完成 (斯佩科特等2001)。
标记
1 ) 所用设备与试剂
细胞培养瓶、液闪计数器、刮 刷 、低磷酸培养基 (无放射性磷酸盐)、裂解缓冲液[20 mmol/L Tris(p H 7.5)]、 2 mmol/L E D T A 和 0.5% S D S 、 N P -40、 DNase I。
2 ) 操作步骤
(1) 用 60m m 或 I O O m m 培养瓶,在完全培养基中培养细胞至所希望的密度。
⑵用预热的低磷酸培养基 (无放射性磷酸盐) 冲洗 2 次 ,然后加人含 0.5〜ImCi32 P/ml(ICi = 3.7xl010 Bq) 的低磷酸培养基 (50——100umol/L 无机憐酸)。
(3) 培养结束后,回 收 32 P 标记培养基,用 IOml Aquasol(New England Nuclear) 稀 释 ,取一等份用液闪计数器测定 32P 活性。测定培养基中磷酸浓度,从这两个数据计算标记培养基中 32P 比活性。
⑷ 用 无 二 价 离 子 的 P B S 清 洗 32P 标记的细胞2 次。每 个 I O O m m 培 养 皿 加 Iml 预热的裂解缓冲液在沸水浴中裂解细胞。用一次性细胞刮刷将细胞裂解物刮到一边,然后吸到 1.5m l 离心管,热 水 浴 l 〇 min。
(5) 冷却至室温。
(6) 离心留上清,加 仰 - 4 0 至 2 % ,加 MgCl2至5mmol/L。加无蛋白酶的DNaseⅠ 与RNase 各 0.I m g ,室温放置 20 min
3 ) 注意事项
(1) 标准组织培养基有 l——2 mmol/L 磷酸盐。为 提 高 32P 标记效率,通常将培养基中磷酸盐浓度降至 5 〇 umol/L 或者更低。标记时间决定于胞内 A T P 库与培养 基 中 32 P 比活性达到平衡的快慢。
(2) 对于长时间培养,为避免类似成纤维细胞这样的快速分裂细胞生长减慢,培养基中无机磷酸浓度应保持在 50〜 100 mmol/L 。
(3) 测定培养基中磷酸浓度时也可用高效液相层析从其他 32P 标记的分子中分离A T P ,直接测胞内 [Y-32P] A T P 的比活性。
⑷ 2——4 h 的短脉冲标记可以用来研究瞬时的蛋白质磷酸化。然 而 ,在脉冲标记实验中 ,蛋 白 质 32 P 标记的暂时提高,可能是磷酸化的提高,也可能因为磷酸转换增强,而蛋白质稱联的磷酸的量并没有改变。分别用含有或不含刺激物的样品,用双向凝胶电泳/W estern 印迹比较未磷酸化蛋白质对磷酸化酸性同工型的比率,就可以区别以上两种可
能性。
检 测 32 P 标记的蛋白质
放射性磷酸 32 P 发出高能 P 粒子。因此,可 以 用 X 射线片放射自显影检测在单向或者双向凝胶上分辨出来的32 P 标记的蛋白质。
1 ) 所用设备与试剂
Whatman 3M M 滤纸、液闪计数器、胶片。
2 ) 操作步骤
(1) 单向或双向凝胶电泳分辨 32 P 标记的样品。
(2) 在 Whatman 3M M 滤纸上吸干凝胶。用放射性墨水在凝胶边缘标几个点。
(3) 在完全黑暗或安全光下,在 X 射线片曝光夹里,将胶片放在凝胶与增感屏之间。确保胶片与凝胶增感屏的闪烁体面紧密接触。让凝胶与胶片夹在两块增感屏之间。用两块塑料板夹紧曝光夹,于- 7 0℃ 曝光。
(4) 冲 洗 X 射线片,显影。
(5) 依放射性记号,在读片机里将考马斯亮蓝染色胶与放射自显影图排列好,标出靶蛋白,拍照。
(6) 切下含祀蛋白的胶条,用液闪计数器检测磷酸化蛋白的放射性。
3 ) 注意事项
放射自显影光密度分析也可以测定蛋白质带的比活性。但是,基于放射自显影对放射性进行精确的定量,需要合适的曝光时间,而且受到胶片狭窄的线性反应期的限制。如 果 有 P-扫描仪,可以用磷光成像板对32 P 放射性进行定量测定。
蛋 白 质 相 互 作 用
基因是相对静态的,而基因编码的产物—蛋白质,贝 U 是动态的。具有时空性和可
215
2 ) 实验步骤
(1) 将细胞裂解液与 5 0ul 的 5 0 % 谷胱甘肽琼脂糖球珠悬液和 25 飓 G S T 在 4℃ 颠倒混合 解 育 2 h 。
(2) 在微量离心机上以最大速度在 4 ¾ 离心混合物 2 min。
(3) 将上清转移到新的离心管中。
(4) 设定两个含等量上清及 5 0ul 谷胱甘肽琼脂糖球珠的微量离心管。在一管中加约10 ug 的 G S T 融合探针蛋白。两个反应中加入的探针和对照蛋白质的量应当是等摩尔的。将离心管在 4℃ 颠倒混合孵育 2 h。
(5) 在微量离心机上以最大速度离心样品 2 min。
(6) 在新的微量离心管中收集上清。这样样品可通过步骤 (10) 中 的 S D S -P A G E 进行分析。
(7) 用 Iml 冰冷的裂解液洗涤球珠。在微量离心机上以最大速度离心lmin。弃去上清 。重 复 洗 3 次 。
(8) (可选) 加人 50ul 20 mmol/L 的还原型谷胱甘肽 (在 50 mmol/L Tris • Cl p H 8.0 中) 到球珠中,洗 脱 G S T 融合蛋白及任何与其结合的蛋白质。在微量离心机上以最大速度离心2 min
(9) 将球珠 [来自步骤 (7)] 或洗脱蛋白质 [来自步骤 (8)] 与 等 体 积 S D S -P A G E 上样缓冲液混合。
(10) 将样品煮沸 4 min 并 作 S D S -P A G E 分析。
3 ) 结果判定
检 测 与 G S T 融合蛋白结合的蛋白质的方法取决于细胞裂解液是否被 35 S 标记以及试
验目的。如果目的是检测与融合蛋白结合的所有35 S标记的蛋白质,就在干胶仪上将胶抽 干 ,并 用 X 射线片曝光进行放射自显影。如果目的是检测特异性结合的蛋白质,就将蛋 白 质 从 S D S 聚丙烯酰胺凝胶转移至膜上,进行免疫印迹分析。如果目的是确定与融合蛋白结合的蛋白质的大小和丰度,而这些蛋白质是来自非放射性的裂解液,就用考马斯
亮蓝或硝酸银将胶染色。
免 疫 共 沉 淀
免疫共沉淀是以抗原和抗体之间的专一性作用为基础的研究蛋白质间相互作用的经典方法之一。在细胞的非变性条件下被裂解时,完整细胞内存在的许多蛋白质-蛋白质结合被保持下来。如果用蛋白质的抗体免疫沉淀该蛋白质,那么与该蛋白质在体内结合的另一蛋白质可能也沉淀下来。基于与蛋白质的生理性相互作用,蛋白质的免疫沉淀归类为免疫共沉淀。这种方法最常用于测定两种目标蛋白质是否在体内结合,但也可用于确定一种特定蛋白质的新的作用搭档。在保持蛋白质与蛋白质相互作用的条件下,收获和裂解细胞,从细胞提取液中特异性地免疫沉淀目的蛋白质,然后通过聚丙烯酰胺凝胶电泳分离免疫沉淀物。该方法的优点是相互作用的蛋白质都是经过翻译后修饰的,处于天然状态,蛋白质相互作用可以在自然状态下进行,能避免人为的影响,分离到天然状态的相互作用蛋白质 (Kay 2003)
217-221
水 和 大 约 3 0 粒无菌玻璃珠。② 把 5 个平板叠放在一起,紧握平板,猛烈地摇动平板直到菌落重新悬浮。③用无菌移液管从每个平板中吸取 5m l 酵 悬 液 。把细胞悬液集中在 50m l 的无菌锥形瓶中。④继续下面 5 个平板,重复①-③ I连 续 从 3 0 个平板中收获酵母细胞,结果得到总体积为 150m l 的酵母悬液,保 存 在 3 个 50m l 的瓶中。
ii.如果需要, 在每个装有酵母细胞的锥形瓶中加无菌 T E (p H 7.5 域无菌水至4〇 〜45m l ,振荡或翻转瓶中悬浮细胞。
iii. 使用台式离心机,室温、 1000——1500 g 离 心 5 min,弃去上清。
iv. 重 复 ii 和 iii。
v. 重新悬浮压紧的细胞沉淀于1 倍体积的无菌甘油溶液中,合并不同管的内容物 ,彻底混合。
Vi.分别转移 Iml 的细胞混合物于一系列无菌小离心管中, - 7 0℃ 冻存。
c.相互作用蛋白的筛选:
i. 解冻一份转化文库酵母细胞,用 C M (Gal-Raff)-Ura-His-Trp 培 养 基 按 1 : 10稀 释 , 3 0 ℃ 摇动培养酵母细胞 4 h 来诱导文库中 G A L l 启动子的转录。
ii.在适当数量的 100m m C M (Gal-Raff)-Ura-His-Trp-Leudropout 平板上分别培养 IO6 个细胞。
iii. 30℃ :培养平板 5 天。
iv.观察平板的生长,出现克隆时对其加以标记。
V. 第 5 天,涂布一^按^天出现的不同克隆分组的主平板 C M (Gal-Raff)-Ura-His_Tip。
vi. 3 0℃ 孵育平板直到菌落出现。
d. 阳性相互作用的确定:弟半乳糖苷酶活性和亮氨酸需求的检测。
e. 测定转录活化:①使用一个平头牙签分别在下面的 4 个平板上复制和主平板上相同的栅格。使用相同的牙签在 4 个平板上划线培养单克隆,尽量在含有 X-gal平板上划制粗的酵母线条,而在亮氨酸缺乏的平板上划细的酵母线条。②平板在 30℃:培 养 3〜 4 天 。
f.解释结果。如果具备下面的条件,菌落和它们包含的质粒被认为第一轮的阳性克 隆 :在 C M (Gal)-Ura-His-Trp 平板上的培养物 X -gal 呈现蓝色;在 C M (Glu)-U m -His-Trp 平板上的培养物 X -gal 分析呈现白色或仅是浅蓝色; 菌落在 C M (GalRaff)-Ura-His-Trp-Leu 平板上生长良好;菌落在 C M (Glu)-Ura-His-Trp-Leu 平板上生长缓慢或根本不能生长。
噬 菌 体 展 示 技 术
噬菌体展示技术是将表达多肽的基因与噬菌体表面蛋白编码的基因融合,以融合蛋白的形式表达在噬菌体表面的一种方法,最常用的表达系统是 M 1 3 噬菌体。将 c D N A 文库插入噬菌体载体进行表达后,所得到的总体为噬菌体展示库。为 了 得到与「诱 饵 」蛋白 相 作 用 的 「猎物」蛋白,可将展示库与固定化的「诱馆」蛋白相互作用,则 「猎 物 」蛋白被吸附下来,被吸附的重组噬菌体可经再感染而扩增,从而得到猎物蛋白的插入基
因 ,目前,噬菌体展示技术可用于鉴定和分析与其他蛋白质相互作用的蛋白质的表面特征 ;分离与特定氨基酸序列结合的新的配体;提高配体与靶结构之间相互作用的亲和力和特异性 (Brymora 2004)。
在微量滴定板上进行亲和筛选的方法,是将标靶蛋白固定在微量滴定板上,噬菌体重组肽库加到板上来选择相互作用的蛋白质。淘选过程可以再重复 2 次 ,以富集可结合的噬菌体减少非特异性结合噬菌体的回收。经 过 3 次的淘选,特异性连接的单克隆可通过 E L I S A 进行检测。连接上的可以在加入辣根过氧化物酶标记的抗噬菌体单克隆抗体及显色底物后呈现颜色。
噬菌体表面呈现库的构建主要包括以下的步骤:
(1) 首先对要呈现的目的基因片段进行随机酶切,并对酶切片段进行回收;
(2) 酶切片段和克隆接头进行连接,并插入噬菌体载体;
(3) 重组的噬菌体转人合适的宿主菌建库;
(4) 转化菌培养扩增后提取重组的噬菌体。
试剂与仪器
(I) 0.2mol/L 碳酸氢盐缓冲液 (Na2 H C O 3), p H 8.5;
⑵ 封 闭 缓 冲 液 :含 有 l%(m/V) BSA 的 PBS;
⑶ 洗 脱 缓 冲 液 :含 有 0.05%(W V )Tween-2 〇的 P B S ;
(4) 重组噬菌体展示库;
(5) 酸 洗 脱 缓 冲 液 : 50 mmol/L 甘氨酸-H C 1, p H 2.0;
( 6 ) 中和缓冲液: 0.2mol/LTris • Cl, pH7.5;
(7) 细菌: D H 5a F 培养过夜;
(8) 2x Y T 培养基以及顶层、底层琼脂:
胰蛋白膝 IOg
酵母膏 IOg
NaCl 5 g
溶 于 I L 的水中,高压灭菌。 4 ℃储 存 2 周 ;
(9) 底层琼脂:高压前加琼脂至终浓度为 1.5% (m/V);
(10) 顶层琼脂:高压前加琼脂至终浓度为0.8% (m /V);
(11) 阴性对照蛋白;
(12) 辣根过氧化物酶标记的抗噬菌体 M1 3 的单克隆抗体;
(13) 显色底物;
(14) 9 6 孔微量滴定板;
(15) 移液管;
(16) 5m l无菌管;
(17) 无菌牙签;
.
(18) 能读取微量滴定板的分光光度计;
(19) DNA 纯化测序设备。
方法
1 ) 微量滴定板包被
⑴ 纯 化 的 目 的 蛋 白 1〜 3 ug加 入 〇.5m l 的 0.2mol/L 的碳酸盐缓冲液 (p H 8.5 冲 混 合9 6 孔板中每孔加 1500,在室温下静置孵育 2 h ,少 数 蛋 白 质 可 以 代 过 夜 。
(2) 每孔加入 2 〇〇ul 的封闭缓冲液室温孵育 3 〇 min。
(3) 轻轻倒掉液体,在纸上轻叩两下,去除残留溶液。
(4) 每孔中加人 200叫洗脱缓冲液,洗 孔 3 次。
2) 选择相互作用的嗔菌体
⑴稀释重组噬菌体展示库,每 150ul 洗脱缓冲液含 1000 个噬菌体,之 后 取 150ul加入到孔中,室温孵育 2 h。
(2) 轻轻倒掉液体,在纸上叩干,去除残留溶液。
(3) 加 人 200|il 洗脱缓冲液,洗 孔 3 次。
3 ) 结合嗔菌体的回收
(1) 加 人 150ul 酸洗脱液,洗脱,在室温放置 lOmin,将 3 个孔的液体收集到一个干净 的 5m l 的塑料管中,每孔含有 1〇 3 ——1〇 8 个噬菌体颗粒。噬菌体也可以用〇.2m o l/ L 乙醇胺 (PH12.0) 洗 漆 ,然后中和。
(2) 加 人 200ul过夜培养的细菌,室温孵育 l〇 min。
(3) 加 人 3 ml 2x Y T 顶层琼脂培养基,混勻,倒 人 100个 15m m 的含有 2x Y T 底层琼脂培养基的培养皿中,在室温放置 5 min 凝 固 ,然后将板放于 3 7 ¾ 孵育过夜。(除了用板扩大噬菌体外,还可在液体培养基中扩大。将感染后的细菌的溶液加入到 5m l 的 2x Y T的肉汤中,转 移 至 50m l 的管中,于 37℃ 摇动过夜。)
⑷ 往 顶 层 琼 脂 上 加 人 5m l 洗脱缓冲液,用于洗提噬菌体颗粒,代 静 置 过 夜 。
(5) 取 5 0 0ul 噬菌体悬液13 200r/m i n 离 心 IOmin, 将上清移入新管中,这样就得到了噬菌体。加 400 ul 20%(m /V)P E G /2.5mol/LNaCl 到 1.6 ml 的上清中,冰浴 15 min,迅速
以 13 200r/m i n 离 心 5 min,再 用 2 0 0 0 洗脱缓冲液重悬。
( 6 ) 重复上述过程,以获得大量的噬菌体。每个循环收集到的噬菌体滴度为 IxlO11 pfu/ml
( 7 ) 收集到的噬菌体在 PB S 中 以 I : 100 稀释。在无菌的 5m l 的管中加入 200ul 过夜培 养 的 D H 5 〇 iF, 1〜 IOul 稀释后的噬菌体, 3m l 的 2x Y T 顶层琼脂培养,混勻。
(8) 将稀释的噬菌体接种在 2x Y T 底层琼脂培养基上, 3 7 ¾ 孵育过夜。
(9) 用无菌的牙签从噬菌斑挑取噬菌体,加 入 到 含 有 50ul 的过夜培养的 D H 5cxF 及lml2x Y T 培养基中。 37℃ 通风过夜培养生长。
4) ELISA 测定
(1) 9 6 孔板中一个孔用标靶蛋白包被,另一个用阴性对照蛋白包被,见 「 1) 微量滴定板包被」的步骤 (1)〜 (4)。 注意设置足够的孔用于检测「 3) 结合噬菌体的回收」中的步骤 (9) 得到的克隆。
⑵每对成对的孔中 (目标与对照伽入 1〇〇ul洗脱缓冲液和 2 5ul 的 「3) 结合噬菌体的回收」中的步骤(9) 得到的单克隆噬菌体的上清,室温孵育 2 h。
(3) 轻 轻 倒 掉 液 体 ,在纸上叩干,去除残留溶液。加 入 20〇 ul 洗脱缓冲液,洗孔3 次 。
⑷ 每 孔 加 l〇〇 ul 的抗噬菌体单克隆抗体(连有辣根过氧化物酶),以 1 : 5000 的比例稀 释 ,室 温 孵 育 lh。
(5) 轻轻倒 掉 液 体 ,在纸上叩干,去除残留溶液。加 入 200ul 洗 脱 缓 冲 液 ,洗孔3 次 。
(6) 每 孔 加 入 IOOul的显色底物,孵 育 3 〇 min。
(7) 4〇5n m 处测量光密度值。
5 ) 确 定 相 互 作 用 的 噬 菌 体 序 列
(1) 插入到重组噬菌体中的 D N A 纯化及测序。
(2) 将编码序列转化为肽序列。
噬菌体展示可用于:①鉴定和分析与其他蛋白质相互作用的蛋白质的表面特征;②分离与特定氨基酸序列结合的新配体;③提高配体与靶结构之间相互作用的亲和力和特异性。
蛋 白 质 结 构
蛋白质有成千上万种,不同的蛋白质具有不同的生物功能,生物功能的多样性是由蛋白质结构多样性所决定的。因此,蛋白质结构的研究占相当重要的地位。
蛋 白 质 一 级 结 构 的 测 定 方 法
蛋白质一级结构的测定有助于研究蛋白质的立体结构和蛋白质的功能,了解物种分子进化及鉴定分子病等。
1 ) 蛋 白 质 样 品 的 纯 化
蛋 白 质 样 品 与 等 体 积 S D S - P A G E 上 样 缓 冲 液 混 合 。将 样 品 煮 沸 4m i n 后进行SDS-PAGE 电泳。
2) 测 定 蛋 白 质 分 子 中 的 多 肽 链 的 数 目
通过测定蛋白质分子中 N 端 和 C 端残基的摩尔数及蛋白质的相对分子质量,可以确定蛋白质分子中肽链的数目,但应注意可能有封闭的末端,如乙酰化的 N 端和酰胺化 C端等。
3 ) 多肽链的分离
在测定一个蛋白质的结构以前,首先必须保证被测蛋白质的纯度,使结果准确可靠。其次要了解它的分子质量和亚基数,按照其亚基数将蛋白质分成几个多肽链。
(1) 肽链的拆开。蛋白质分子多肽链的连接有共价结合和非共价结合两种。要拆开以共价结合的一 S- S—连接的多肽链,必须采用的化学处理方法有:
a.过甲酸氧化:用氧化剂过甲酸断裂一 S—S— 。这个反应一般在〇丈下进行 2 h左 右 , 2 个 S 就全部能转变成磺酸基,这样被氧化的半胱氨酸称为磺基丙氨酸。
如果蛋白质分子中同时存在半胱胺酸,那么也会被氧化成磺基丙氨酸。此外甲硫氨酸和色氨酸也可被氧化,从而增加分析的复杂性。
b. 巯基乙醇还原:利用还原剂巯基乙醇亦可使蛋白质的一 S_s 一断裂。当高浓度的巯基乙醇在 P H 8.0 条件下室温保温几小时后,可以使一 s s 一定量还原。与此同时反应系统中还需要有 8mol/L 脲 或 6mol/L 盐酸胍使蛋白质变性,多肽链松散成为无规则的构型,此时还原剂就可作用于一 S—S— 。此反应是可逆的,
因此要使反应完全,巯基乙醇的浓度必须在 0.1——0.5moi/L。
c. Cldand 试剂的还原作用 Cleland 指出二硫赤藓糖醇 (dithioerythritol) 及二硫苏糖醇 (dithiothreitol)在氧化还原能力上是比较强的试剂,只 要 0.01mol /L 就能使蛋白质的一 S- S—还原,反应基本与巯基乙醇相似,且在许多球蛋白反应中,可以不用变性剂。 Cldand 试剂首先与蛋白质一 S- S—形成中间物,反应终了,还原剂被氧化形成一个稳定的六环化合物,蛋白质则被还原。蛋白质分子的几条肽链若以非共价键结合,则用尿素、盐酸胍等变性剂即可拆开。
(2) 蛋白质的多肽链被拆开后,将它分离纯化,一般多用凝胶过滤、离子交换、电泳等方法。
4 ) 测 定 多 肽 链 的 氨 基 酸 组 成
(1) 代 测 蛋 白 质 与 5.7 mmol/L 的盐酸在水解管中混合。并加入苯酚以防半胱氨酸和甲硫氨酸氧化,抽去空气,充满氮气, 110℃ 水 解 24 h ,或 在 150℃ 下快速水解 90 min。
(2) 水解得到的氨基酸与化学试剂反应,产生易被检测的氨基酸衍生物。氨基酸与茚三酮反应生成紫色化合物,在 570n m 处有吸收峰。脯氨酸和羟脯氨酸与茚三酮反应生成黄色化合物。
(3) 分离及检测。酸性条件下氨基酸带正电荷,是阳离子。在通过阳离子交换树脂时被吸附到柱上,再用洗脱液洗脱、收集。
5) N 端 和 C 端 氨 基 酸 残 基 的 测 定
N 端分析:①多肽的 a-氨基与异硫氰酸苯酯 (PITC) 试剂耦联;②在无水条件下用三氟乙酸酸裂解,形 成 A T Z -氨基酸;③ A T Z -氨基酸在三氟乙酸水溶液(25%) 中转化成稳定的 P T H -氨基酸;④通过薄层层析、气相色谱、高效液相谱及质谱进行氨基酸的鉴定。 C端分析: C 端自由羧基与乙酸酐反应生成混合酸酐,并进一步环化成曝唑酮, C 端® 唑酮在硫氰四丁胺的作用下,转化为乙内酰硫脲 (T H )。
(1) N 端测定:
a. 二硝基氟苯法(F D N B 、 D N F B ): D N P -氨基酸用有机溶剂抽提后,通过层析位置可鉴定它是何种氨基酸。 Sanger 用此方法测定了胰岛素的 N 端分别为甘氨酸及苯丙氨酸。
b. 氰酸盐法。
c. 二甲基氨基萘磺酰氯法。
(2) C 端分析:
a. 肼解法:这 是 测 定 C 端最常用的方法。将多肽溶于无水肼中, 1 0 0 ℃下进行反应 ,结果竣基末端氨基酸以游离氨基酸状释放,而其余肽链部分与肼生成氨基酸肼。这样羧基末端氨基酸可以采用抽提或离子交换层析的方法将其分出而进行分析。如果竣基末端氨基酸侧链是带有酰胺如天冬酰胺和谷氨酰胺,则肼解时不能产生游离的羧基末端氨基酸。此外肼解时注意避免任何少量的水解,以免释出的氨基酸混淆末端分析。
b. 羧肽酶水解法:羧肽酶可以专一性水解羧基末端氨基酸。根据酶解的专一性不同,可区分为羧肽酶 A 、 B 和 C 。应用羧肽酶测定末端时,需要事先进行酶的动力学实验,以便选择合适的酶浓度及反应时间,使释放出的氨基酸主要是 C端氨基酸。
6 ) 肽 段 的 氨 基 酸 序 列 分 析
E d m a n 降解的最大优越性是在水解除去末端标记的氨基酸残基时,不会破坏余下的多肽链。
⑴ 用 苯 异 硫 氰 酸 酯 (PITC)在 p H 9.0 的碱性条件下对蛋白质或多肽进行处理, PITC与肽链的N 端的氨基酸残基反应,形成苯氨基硫甲酰 (P T C ) 衍生物,即 F T C 肽 。
(2) P T C -肽用三氟乙酸处理, N 端氨基酸残基肽键被有选择地切断,释放出该氨基酸残基的噻唑啉酮苯胺衍生物。
⑶将该衍生物用有机溶剂(如氯丁烷) 从反应液中萃取出来,而去掉了一个 N 端氨基
酸残基的肽仍留在溶液中。萃取出来的噻唑啉酮苯胺衍生物不稳定,经酸作用,再进一步环化,形成一个稳定的苯乙内酰硫脲 (P T H ) 衍生物,即 P T H -氨基酸。留在溶液中的减少了一个氨基酸残基的肽再重复进行上述反应过程,整个测序过程现在都是通过测序仪自动进行。每一循环都获得一个 P T H -氨基酸,经 H P L C 可以鉴定出是哪一种氨基酸。当蛋白质中含有一个或多个半胱氨酸残基时,有时一对半胱氨酸残基会通过二硫键交联。在这种情况下测序,首先要经过处理 (如用过甲酸处理) 切断二硫键,然 后 再 进 行 Edman降解测。
7 ) 重 叠 法 确 定 肽 链 的 一 级 结 构
8 ) 蛋 白 质 分 子 中 二 硫 键 及 酰 胺 键 的 确 定 和 磷 酸
化 、糖 基 化 修 饰 点 的 定 位
(1) 蛋白质分子中二硫键的定位。
a. 蛋白质中由二硫键相连的肽链拆分:
i. 过甲酸处理:蛋白质中带游离巯基的半胱氨酸残基及由二硫键连接的半胱氨酸残基可被过甲酸氧化成黄基丙氨酸残基,将肽链拆分。
ii. 疏基乙醇或二巯基苏糖醇处理:用巯基乙醇或二巯基苏糖醇还原二硫键,再用碘乙酸烷基化,然后分别测定拆分得到的多肽链的一级结构。
b. 酶水解肽链未拆分的蛋白质,找出其中含有二硫键的肽段:蛋白质在微酸环境中进行 (P H 2.0〜 6.5)。 将肽段样品点样在正方形滤纸一角,第一方向碘用分离后,用过甲酸蒸气处理滤纸,然后滤纸旋转 90°,在与第一方向电泳的条件下进行第二方向电泳。由于两次电泳条件相同,不含二硫键的肽段在两次电泳中迁移率相同,最终聚集在正方形滤纸的对角线上。含有二硫键的肽段,在过甲酸处理后产生两个负电性增强的肽段,第二次电泳迁移率与第一次不同,不聚集在对角线上,据此可以加以区分。
(2) 酰胺基的定位。蛋白质的氨基酸组成分析可测定蛋白质中酰胺氮总量。
(3) 磷酸化修饰定位。用 ESI 质谱法或 MALDI 质谱法比较憐酸化前、后肽段的质谱可以定位磷酸化的位点。
(4) 糖基化位点的确定。肽段中糖计划修饰有两种类型,N-糖基化发生在门冬酰胺的侧链 N 原子上, O -糖基化发生在丝、苏氨酸侧链羟基的〇原子上,比较用糖苷酶水解前 、后糖肽的M A L D I M 谱便可以确定修饰位点 (鲁 子 贤 1982)。
蛋 白 质 二 级 结 构 的 测 定
1 ) 圆二色谱分析
圆二色谱分析是检测蛋白质二级结构信息的一种有用的方法。不同的二级结构对应与左旋光和右旋光的光吸收差值。通过分析圆二色谱曲线,可以估计二级结构螺旋、折叠以及无规则卷曲的比值。
样品的处理: Res0urseQ 和 R esourseS 离子交换柱分离的样品,低温中速离心浓缩至0.2 mg/ml,并将缓冲液换至测量缓冲液(浓度为 5 〇 m mol/LTris、 50 mmol/LNaCl、 5 mmol/LMgCl2、 p H 7.5 的缓冲液中含 0. 2 m g/mi) 中,直接用于测量。测 量 杯 长 度 为 0.2 cm , 溶液体 积 l 〇〇 ul。波长频率范围是 190〜 250nm。测量温度为 2 5 ℃
2 ) 动态光散射分析
动态光散射仪 (dynamic light scattering) 分 析 ,在 Protein Solution 公司的 Dyna Pro 仪器上运行。
样品处理: ResourseQ 和 ResourseS 离子交换柱分离的样品,低温中速离心浓缩至0.2 mg/ml,并将缓冲液换至测量缓冲液(浓 度 为 50 mm〇l/L Tris、 50 mmol/L NaCl、 5 mmol/LMgCl2、 pH7.5 的缓冲液中含〇.2 mg/ml 样品)中,取 2〇ul 蛋白质样品, 12 000r/min 高速
离 心 IOmin 后 ,用于测量。在 2 0 次的测量后,计算出溶液状态中的蛋白质的分子半径、分子质量以及主要的颗粒。测量温度为 25 丈 (Wei 1996)。
蛋 白 质 晶 体 空 间 结 构
X 射线衍射法分析蛋白质的晶体空间结构。
1 ) 材料与仪器
R I G U K U 转靶阳极 X 射线发生器安低温液氮控制系统、衍射数据收集的面探测仪、X 射线单色器。
2 ) 方法
X 射线晶体学研究通常采用的 X 射线波长与化学键键长相当,也与晶体内的原子间距离响应,约 为 O.lnm。一个晶体包括上亿个有序排列的基本单元。在晶胞中,原子核外电子组成的电子云对 X 射线产生衍射,由于晶体的周期性衍射束只在特定方向出现,电子云可以用傅里叶级数展开,其中每一项的大小与衍射强度相对应,可直接测量,而每一项的相位目前尚没有成熟的方法直接进行测量(牛 立 文 1998)。
(1) 结 晶 :需要通过大量的条件筛选和优化以恰好是蛋白质分子间的相互作用促成蛋白质分子形成高度有序的晶体,而不是随机聚合形成沉淀。这就要求溶液中的蛋白质处于合适的过饱和状态,并只形成少数的成核中心,使晶体能持续地慢慢生长成大单晶。
(2) 数据收集:通常利用 (单波长)X 射线光束照射在一定角度范围内旋转的蛋白质晶 体 ,同时记录晶体对 X 射线散射的强度。这些强度可转换为结构测定中的结构因子的振幅。
(3)相 角 的 测 定 :结构因子的振幅及相角不是直接相关的量。由于结构因子相角的全部信息在收集数据时丢失,因此必须通过其他途径来得到它们的数值。
(4)相角的改进 (优化):电子密度图的质量及其后的可解释性主要取决于相角的准确性 。有的情况下采用晶胞中不对称单元中的等同部分的电子密度平均,有可能大大地改善误差较大的起始相角。
(5) 电子密度图的解释:相位确定后,可开始计算电子密度图。若从电子密度图能跟踪出肽链走向和分辨出二级结构,则可能推出多肽链的三维折叠方式。进而根据氨基酸序列,就可能构建出原子坐标形式的蛋白质结构模型。
(6) 修 正 :考虑到以建立的立体化学资料的限制,根 据 X 衍射数据对初始的蛋白质分子模型进行修正 (Kensal 1998)。
###载 体 的 制 备
目的基因的克隆是分子生物学实验中的核心内容,起着承上启下的作用,通过克隆技术可实现目的基因的无限扩繁、长期保存的目的;利用该技术能实现基因-组、基因改造、构建工程菌和转基因动植物。基因克隆技术主要包括载体的获取、酶切、脱憐、连接和转化及筛选、鉴定等内容 (奥斯伯等 2001)。在干细胞增殖、分化调控机制的研究中常用到的克隆载体主要有以下两类:①宿主为大肠杆菌的细菌质粒;②动物细胞为宿主的动物病毒和以动物病毒为基础改造的穿梭质粒。
载体特点:①在宿主细胞中具独立复制能力;②带抗性等选择标记;③有合适的限制性内切核酸酶位点,可插人一定片段长度的外源 DNA 而不影响自身复制。
### 质 粒 提 取
质 粒 D N A 的提取方法比较常见,基本步骤多为三步:①细菌培养和质粒的扩增;②细菌菌体的裂解;③ 质 粒 DNA 的纯化。现在常用的是质粒提取试剂盒。该方法方快 捷 ,具体步骤可参考其使用手册。
注意:在 质 粒 DNA 提取中,质 粒 D N A 的质量和产量在很大程度上受培养条件的影响 ,通过一些实验发现,下列培养方式能获得较好的结果。①培养应用菌龄不超过 4 天的平板单菌落作为起始菌。②培养菌应在 37℃、 220r/min 下 生 长 16—— 18 h。③不能用经储藏的培养菌直接进行质粒提取制备。
### D N A 片 段 的 酶 切
限制性内切核酸酶 (restriction endonuclease) 是一类能识别双链 DNA 分子中特异核苷酸序列的 DNA 水解酶,是体外剪切基因片段的重要工具。
1 ) 主要设备与试剂
离心机、恒温水浴、电泳仪、电泳槽、质粒、 TAE 缓冲液、限制性内切核酸酶
2 ) 操作步骤
用于质粒的酶切鉴定,一 般 是 20ul体积中含有0.2〜Iug D N A , 酶切反应体系为:
质 粒 DNA 10ul
缓冲液 (IOx) 2ul(双酶切时要选取两种酶都适合的缓冲液)
限制性内切核酸酶 1ul(双酶切时两种酶各加 1 ul)
双蒸水 7ul
⑴ 在 E p 管中加入 7 ul双蒸水。
⑵ 加 人 IOx 缓 冲 液 2ul 。
(3) 加 限 制 酶 1 ul。
⑷加入质粒 10 ul
(5) 离 心 ,混匀。
(6) 37℃水浴 l——2 h。
(7) 取 反 应 液 ,电泳鉴定。
3 ) 注意事项
⑴ 样 品 加 入 次 序 为 水 、缓冲液、 D N A ,最后为酶,不应颠倒。
(2) 加酶步骤要在冰浴中进行,在加酶前应先将水、缓冲液及待切 D N A 混匀。
(3) 反应体系的体积要尽量少 , 一 般 水 解 0.2〜 Iug D N A 时 ,应将体积控制在 20——30 ul内。保证限制酶活性不受抑制。
⑷为了控制反应体积和促进反应进行,要求模板 D N A 的浓度应很高,如 底 物 D N A浓度过低则应进行浓缩。
(5) 本实验中双酶切时,因两种酶的缓冲液盐浓度要求相同,所 以 ,可在同—反应体系中完成双酶切。但如进行双酶切时,两种酶所需缓冲液盐浓度要求不同,则不能将两种酶同时加入同一体系进行反应,此时可采取一种酶水解完后,进行有机溶剂抽提,沉淀、干燥后再复溶,然后进行第二种酶切。
### D N A 片 段 回 收
回收是指从电泳介质中纯化出目的片段。在试验中待回收的片段主要有酶切片段和各 种 P C R 片段;回收的介质主要为琼脂糖;回收的方法主要有微量柱回收、玻璃奶回收。这里以玻璃奶法为例介绍 D N A 片段回收的方法。
1 ) 主要设备与试剂
离心机、烘箱、恒温水浴、玻璃奶、无水乙醇、溶胶液、漂洗液。
2 ) 操作步骤
(1) 从琼脂糖凝胶上切下目的条带放入 1.5 ml E p 管中,用 一 个 Tip 头捣碎。
⑵ 准 确 估 计 凝 胶 的 体 积 ,加 入 3 倍体积的溶胶液,室温放置 5m i n 或 50℃:保 温 3 min,其间轻摇 E p 管几次使胶完全溶化
。
⑶ 向 溶 胶 中 加 入 IOpl 玻璃奶,颠倒混勻,冰 浴 lOmin,并且间隔 2〜 3m i n 颠倒混匀一 次 , 12 000r/m i n 离 心 30s,吸弃上清。
(4) 将浓缩漂洗液按 3 : 7 与无水乙醇配成工作液,然后向玻璃奶中加入 2 5 0 ul ,用加样器吸漂洗液轻柔地将玻璃奶冲散混匀, 12 OOOr/m in 离 心 30s ,吸弃上清。
(5) 重复第 (4) 步一次。吸取完漂洗液后再离心 10s,用 Tip 头将最后一点儿漂洗液吸干 净 ,然后放置于 37℃ 温箱中干燥 20 min。
(6) 加 20ul 无菌水,用 Tip 头轻轻混勻, 60T ;水 浴 5 min, 12 OOOr/m i n 离 心 lmin,回收上清即为纯化的 P C R 片段。 -20℃ 储存。
3 ) 注意事项
(1) 玻璃奶使用之前一定要融化并充分混匀。
(2) 干燥后加无菌水后要充分吹打。
###基 因 的 重 组 连 接
重组连接是指将目的基因用 D N A 连接酶连接在合适的质粒载体上,这是基因重组、基因改造的重要中间环节。连接反应可在溶液中进行也可在琼脂块中进行。在本部分仅以重组克隆为目的介绍相应的连接类型。在基因重组克隆中,外 源 D N A 与载体的连接是承上启下的一步操作,根据连接片段末端类型不同,连接方式有黏端连接、平端连接、平黏端连接及单碱基配对的 T 载体连接等。
主 要 设 备 与 试 剂
离心机、恒温设备、 T 4 D N A 连接酶、 10xT4 D N A 连接酶缓冲液。
操 作 步 骤
连接
1) 取干净、灭菌 E p 管,加入下列各试剂
T4 DNA 连接酶缓冲液 1ul
载 体 DNA 片段 1ng
插入目的片段 xng
连接酶 IU(平末端,如为黏端连接,则酶量可小于 1u)
加无菌水至 IOul
X = 目的片段与载体片段的分子比例 X 载 体 n g 数 X 目的片段的碱基数/载 体 n g 数
(目的片段/载体片段的分子比例一般为 3 : 1)
2) 连接反应
2 2 ℃ 保 温 3 h 或 4℃过 夜(Promega 连接酶,黏端连接);
或 22〜 2 6 ℃保 温 Ih (B R L 连接酶,黏端连接);
或 22℃保 温 4—— 16 h (Promega 连接酶,平端连接);
或 14℃ 保 温 16——24 h (B R L 连接酶)。
3 ) 注意事项
(1) 分子生物学实验中主要采用 T 4 D N A 连接酶。
(2) 在分子生物学实验中连接酶主要用于:①基因重组中载体与外源基因的连接;② 接 头 与 D N A 片段的连接,这种连接一般发生在文库构建、分子标记的 A F L P 及 差 ^表达的研究中等。
(3) 其中黏端连接的效率最高,黏端连接与平黏连接中外源 D N A 片段在载体中的插入有方向性。平端连接的效率最低,且载体发生自连的比例较髙,因此在克隆操作中为了克服平连的弊端,通常可通过:① T d T 酶在载体和外源 D N A 片段上转移一互补序列 (核苷酸投影法);②在载体和外源 D N A 片段上加上人工接头,变平连为黏连。
连接产物的转化
通过转化可以实现大量扩增质粒。转化的方法有多种,如电转化法、 CaCl2 法等。一般使用 CaCl2 法 ,从而改变细胞膜的结构,使 外 源 D N A 可以穿过细胞膜进入细胞,然后在选择性培养基中培养转化处理过的细菌,转化成功的细菌可以在抗性培养基上生长形成菌落。
1 ) 主要设备与试剂
摇床、离心机、恒温设备、 L B 培养液、 CaCl2(0.1mol/L )。
2 ) 操作步骤
(1) 从新鲜的平板上挑取一个 D H 5α 单克隆, 37℃摇振培养过夜。
(2) 按 I : 100 的比例将过夜培养的菌液加入新鲜的 L B 培养液中,于 37℃继续培养至 OD<sub>600</sub>为 0.3——0.4。
(3) 将菌液转移到预冷的 50m l 离心管中, 4 ℃ 、: IOOOg 离 心 15m i n 回收菌体。
(4) 倒出上清,将 管 倒 置 Imin 以使最后残留的痕量液体流尽。
(5) 用 IOml 冰预冷的 0.1mol/L CaCl2 重悬每份沉淀,同上离心收集菌体。
(6) 重复步骤⑷。
(7) 每 50m l 初始培养物用 2m l 冰预冷的 0.1mol/L CaCl2 重悬,此即感受态细菌。
(8) 取 200 叫感受态细菌和 D N A (≤10ul, ≤ 5 0 ng 伽入到一个冰冷的聚丙烯管中,轻轻旋转混匀,冰上放置 30 min。
(9) 将管放到 42 ℃水浴中,恰恰放置 90s,其间不要摇动。
(10) 快速将管转移到冰浴中,冷 却 l——2 min。
(11) 每 管 加 800ul L B 培养液, 37 ℃温和摇振培养 60m i n 使细菌复苏,并且表达质粒编码的抗生素抗性标记基因。
(12) 取 2 0 0 0 菌液涂在含 A m p 或 K a n 平板上,将平板置于室温直至液体被吸收,然 后 于 3 7 ℃ 倒置培养过夜。
3 ) 注意事项
(1) 热激温度和时间要严格控制好。
(2) 含 A m p 琼脂平板培养时间不应超过 16 h ,否则容易产生卫星菌落。
⑶ 如 果 是 与 T 载体的连接产物,则将菌液涂布在含 X -gal 和 I P T G 的 A m p 或 Kan平板上。
病毒载体的制备 (详见细胞转染部分)
1 ) 逆转录病毒载体的构建
⑴ 改 造 病 毒 基 因 组 ,切 除 野 生 型 的 娜 、 poZ、 e«v 结构基因,换为目的基因,保留包装信号和 L T R S 序列。
⑵将重组的病毒载体导入包装细胞,包装出有感染能力的逆转录病毒颗粒。
(3) 从包装细胞培养上清中获得包装好的缺陷型病毒,用它转导带有该病毒受体的祀细胞,使目的基因稳定整合至靶细胞染色体。
注意:①插入的目的基因不能超过 7kb , 否则逆转录病毒载体将不能有效包装。②外源基因导入靶细胞的效率很高,通 常 Im l 含病毒的培养基可感染 1 〇 <sub>6</sub>个觀细胞。
2 ) 腺病毒载体的构建 ( ;这部分已经商品化,不做详细介绍)
(1) 构建含外源目的基因的腺病毒穿梭载体。
(2) 制备重组质粒。
(3) 转 染 HEK293 细胞。
(4) 收获含重组腺病毒的上清进行滴度测定和纯化。
(5) 转染靶细胞。
注意: 一般插入外源 DNA 长度不得超过 2kb, 但是通过缺失部分病毒基因组可扩大外源 D N A 的插入容量。
###蛋 白 质 的 制 备
在干细胞增殖、分化调控机制的研究中,常需要进行目的蛋白质的表达。首先我们通过载体制备将目的基因插入表达载体,然后再将该重组载体导入活细胞,以大量表达所需目的蛋白质。
外源基因可以在原核表达,也可以在真核细胞中进行表达。外源基因的原核表达是目的基因在原核细胞中以发酵的方式快速、高效的合成基因产物,常用于原核表达的细胞是大肠杆菌。其特点是:操作简便,而且在较短时间可以实现蛋白质的大量表达,因
此成为科研应用中蛋白质表达的主要宿主。但是大肠杆菌进行蛋白质表达也存在一些缺陷 :①真核蛋白在大肠杆菌中的表达没有得到正确的修饰;②在大肠杆菌中表达蛋白时常形成沉淀成为包含体。
###诱导
常用表达用宿主菌株为 DE3 BL2 1 菌 ,先要准备合适的感受态细胞,取 质 粒 Ing 用无菌水稀释 5 0 倍 ,按前述转化步骤进行操作,筛选出阳性克隆后提质粒并经 D N A 序列测 定 ,确定无误后进行如下操作。
1 ) 主要设备与试剂
摇床、 SDS-PAGE 电泳仪、离心机、 LB 培养基、 IPTG、 Tris • HCI(20mmol/L pH8)、2xloading buffer
2 ) 操作步骤
⑴从新鲜的划线平板中挑取单克隆,接至一定体积的含合适抗生素的 L B 中, 37℃摇床振摇培养至OD<sub>600</sub>为 0.4〜 0.6。
(2) 如果表达载体的原核启动子为 PL 启动子,则 在 30〜 32℃ 培养数小时,使培养液的 OD<sub>600</sub>达 0.4〜 0.6, 迅速使温度升至 4 2 ℃ 继 续 培 养 3——5h。如果表达载体的原核启动子为 ta c 等 ,则 37T 培养细菌数小时达到对数生长期后加 IPTG至终浓度为lmmol/L,继续 培 养 3——5 h。
(3) 取 出 Im l菌液做分析用, 1 〇〇〇g发离心 lmin,沉 淀 加 l 〇〇ul 2xloadingbuffer, IOO℃加 热 3 min, 上 样 进 行 SDS-PAGE。
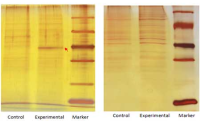
⑷ 将 摇 瓶 置 于 冰 上 5 min。4℃ 、 5 000g 离 心 5m in 收集菌体,重悬细胞于〇.2 5 倍体积预冷的2 〇 mol/L Tris HCl(pH 8.0) 中,离心。菌体保存于-7〇℃冻存以备分离基因表达产物所用。
3 ) 注意事项
不同的表达质粒,因启动子不同,诱导表达方法并不完全相同,可根据具体情况而定。
###细 菌 的 裂 解
细胞破碎是提取胞内产物的关键性步骤。常用的研究方法有酶溶法和超声破碎法。
溶法
酶溶法就是用生物酶将细胞壁和细胞膜消化溶解的方法。常用的溶解酶有溶菌酶(Iysozyme)
1 ) 主要设备与试剂
离心机,裂解缓冲液 (lmmol/L EDTA, 100 mmol/L NaCl,50 mmol/L Tris • Cl,p H 8 . 0),蛋白酶抑制剂 (50 mmol/L ) , 溶菌酶(10m g /ml),脱氧胆酸, D N a SeI(lmg/ml)。
2 ) 操作步骤
(1) 4℃ 、 500 g 离 心 15 min,收集诱导表达的细菌(1000 ml)。
(2) 弃上清液,每克菌加3m l 裂解缓冲液,悬浮沉淀。
(3)每 克 菌 加 3ul 5Ommol/L 的 蛋 白 酶 抑 制 剂 苯 甲 磺 酰 氟 (phenylmethyIsulfonylfluoride, PMSF), 80ul 溶菌酶(10m g /ml),搅拌 20 min。
(4) 在搅拌下,每克菌加 4m g 脱氧胆酸。
(5) 溶 液 变 得 黏 稠 时 每 克 菌 加 20ul DNaseI(lmg/ml)。
(6)室温放置至溶液不再黏稠。
3 ) 注意事项
步骤 (2)〜 (4) 在冷室中进行。
超声破碎法
声 频 高 于 15〜 20kHz的超声波在高强度声能输入下可以进行细胞破碎,在处理少量样品时操作简便,液体量损失少,同时还可对染色体 D N A 进行剪切,大大地降低了液体的黏度,有利于后续目标产物的分离。
1 ) 主要设备与试剂
离心机、 SIDS-PAGE 电泳仪、 T E buffer、 2xloading buffer。
2 ) 操作步骤
⑴收集诱导表达的细菌(IOOOml),4℃ 、 500 g 离 心 15 min。
⑵ 弃 上 清 液 ,每克菌加3 ml T E buffer。
(3) 根据厂家提供的超声波仪的数据进行破菌。
⑷ 10 000g 离 心 15 min, 分别收集上清液和沉淀。
(5) 取 少 量 上 清 液 和 沉 淀 加 入 等 体 积 的 2xloading b u ffe r, 进 行 SDS-PAGE电泳。
3 ) 注意事项
(1) 超声破碎与声频、处理时间、细胞浓度、菌种类型等许多因素有关,实验时要根据具体情况掌握。
(2) 超声波破菌前,细胞经数次冻溶后更容易破碎。
包 含 体 的 分 离 、 溶 解 和 复 性
在以大肠杆菌为宿主菌高效表达外源基因时,表达蛋白常常在细胞质内聚集,形成包含体 (inclusion body)。包含体形成后,表达蛋白不具有物理活性,因此必须溶解包含体对表达蛋白进行复性。由于包含体是致密的,易于沉淀,所以能用脱氧胆酸盐洗涤得到纯度较高的制备物。包含体离心沉淀后,可用尿素等洗涤。为获得可溶性的活性蛋白,必须要将洗涤过的包含体溶解,然后进行重折叠,最后用离子交换或亲和层析的方法对重折叠后的蛋白质进行精制。
分离
1 ) 主要设备与试剂
离心机、 SDS-PAGE 电泳仪、尿素、 Tris . ClCaimolZL, p H 8.5)、 2xloading buffer。
2 ) 操作步骤
(1)4℃,12 OOOg离心细菌裂解液 15 min。
(2) 弃上清液,每 克 菌 加 Iml 无菌水使悬浮,分 别 取 1 0 0 ul 分 装 4 个 管 ,其余备用。
⑶ 离 心 同 步 骤 (1)。
(4) 弃上清液,用 lOOul O.lmolTLTris • Cl(p H 8.5),含不同浓度尿素(0.5mol/L 、lmol/L )的溶液悬浮沉淀。
(5)离 心 。
(6) 分别吸出并保留上清液,用 100ul H2O 重新悬浮沉淀。
(7) 分别取10 叫上清液和重新悬浮的沉淀,加 10ul 2xloading buffer进 行 SDS-PAGE电泳。
包含体的溶解
1 ) 主要设备与试剂
SDS-PAGE 电泳仪,裂解缓冲液 (含 0.1 m m 〇 l/L P M S F ,8mol/L 尿 素 ,10 mmol/L D T T ),K H 2P O 4(50 mmoiyL, p H 10.7), E D T A (lmmol/L , p H 8.0), NaCl(50 mmol/L )
2 ) 操作步骤
(1) 用 IOOul裂解缓冲液溶解包含体。
(2) 室 温 放 置 1h
(3) 加 9 倍体积的 P A G E 电泳缓冲液,室温放置30 min, 用 K O H 保 持 p H 为 10.7。
⑷ 用 H C l 调 至 p H 8.0,最少在室温放置30 min。
(5) 室温、 10 000g离心15min。
(6) 吸出上清液并保存,用 100 ul IxSDS-Ioading buffer重新溶解沉淀。
(7) 取 10 ul 上 清 液 ,加 10ul 2xloading buffer,取20u重新溶解的沉淀,进 行SDS-P A G E 电泳。
复性
1 ) 主要设备与试剂
2 mmol/L 还原型谷胱甘肽和0.2 mmol/L 氧化型谷胱甘肽。
2 ) 操作步骤
(1)每毫升脲-蛋白溶解液缓慢加入9m l 含 2mmol/L 还原型谷胱甘肽和0.2 mmol/L 氧化型谷胱甘肽的工作缓冲液。
⑵ 室 温 孵 育 2〜 4 h 。
融 合 蛋 白 的 纯 化 (以 G S T 标 签 融 合 蛋 白 的 纯 化 为 例)
1 ) 主要设备与试剂
摇床、离心机、超声波发生器、 L B 培养基、 IPTG(100 mmol/L )、 TritonX-100(10%)、5 0 % 的谷胱甘肽-琼脂糖珠、还原型谷胱甘肽洗脱液。
2 ) 操作步骤
(1)挑取一个菌落接种于 100 ml L B /A m p 培养基中,在 3 7 ℃ 摇 床 中 培 养 12〜 15 h。
⑵ 以 I : 1 0 的比例将此培养物稀释于 I L 新鲜的抗性L B 培养基中,分 入 2 个 2L的瓶中, 37℃下 培 养 lh。
(3)加lOOmmol/LIPTG 至终浓度 0.1 mmol/L ,继续培养 3〜 7 h 。
⑷ 室 温 、5OOOg离 心 IOmin收 集 细 胞 ,弃 上清。沉 淀 重 悬 于 10——20m l 冰 浴 的 PBS中。
(5) 将离心管置于冰浴中,用带直径5m m 探头的超声波发生器裂解细胞。
(6) 加 入 1 0 % 的 Triton X -100至浓 度 为 1 % 并混匀。室温、 10 OOOg离 心 5m i n 去除未裂解的细胞。
(7) 上 清 加 入 Iml 5 0 % 的谷胱甘肽-琼脂糖珠混悬液,室温下轻柔混勻2 min。加 50 ml冰 浴 的 P B S 洗涤液、混 勻 , 5 ( % 离 心 10s。重复 洗 涤 2 次 。用少量 (1〜 2 ml)冰 冷 的 PBS重悬琼脂糖珠,并 移 人一个1.5m l 微量离心管中。
(8) 室温、 5O O g 离 心 IOs收集琼脂糖珠,弃上清。加 Iml还原型谷胱甘肽洗脱融合蛋白质。轻 柔 混 勻 2 min, 500 g 离 心 10s,收集上清,重 复 洗 脱 2——3次 ,经 SDS-PAGE分析各部收集样品。洗脱的蛋白质分成小份,加 甘 油 至 终 浓 度 1 0 % ,储存于-70℃。同时测量吸光度确定融合蛋白的产量。
###抗 体 的 制 备
一般来说,抗体制备主要分为两种!一种是用纯化的抗原免疫动物,获得多克隆抗体 ,这种方法在试验中比较常用;另一种是用杂交瘤技术制备单克隆抗体。
多 克 隆 抗 体 的 制 备
一般用纯化抗原免疫动物,可以得到特异性抗血清。免疫动物的抗原虽然进行了纯化 ,可每种抗原分子常有多个抗原决定簇,可以刺激动物产生多种质量不同的抗体,因此异种抗血清是由多种抗体组成的混合物,这类抗血清称为多克隆抗体,在实验研究中应用较广。我们以常见的蛋白质抗原免疫家兔制备抗血清为例进行介绍。
1 ) 所用设备与试剂
注射器、眼科剪、止血钳、引流管、无菌的玻璃瓶、家兔、弗氏完全佐剂、麻醉剂、叠氮钠(N a N 30.02%)。
2 ) 操作步骤
(1) 选择动物: 2.5〜 3k g 的健康家兔。
(2) 免疫动物:基础免疫所需抗原量为 0.5〜 lmg/kg,与弗氏完全佐剂以等体积乳化后做双后足掌注射及背部皮内多点注射。加强免疫,在第一次注射后1 〇天,用前次相同的抗原量与弗氏完全或不完全佐剂乳化,做腋窝淋巴结内注射,每 侧 a i ——〇.2m l ,剩余量做背部皮内多点注射, 1〜2周后,再重复注射一次可不加佐剂,抗原剂量适当增加,注射途径可多点皮下、肌肉或静脉内加强免疫。以 后 每 隔 1——2周 注 射 2——3次 ,末次注射后7——10 天试血,如果抗体效价不高,可再继续加强,直至达到要求的抗体效价(以双向扩散法检测,抗 体 效 价 在 1 : 3 2 以上),方可采血。
(3) 采 血 :颈动脉放血法。麻醉后将兔仰卧,四肢固定,充分暴露颈部,剪 毛 ,消毒 ,沿气管表面做皮肤直切口,暴露气管,在气管旁剥离动脉与神经束。用止血钳夹住动脉的近心端,用丝线结扎远心端,用眼科剪在两端被阻断的血管上剪一 「V 」形小口,向心插入引流管用丝线将插管与血管固定,放开近心端管钳,收集血液于清洁无菌的玻璃瓶中,置室温,待血块收缩完全后分离血清,加 最 终 浓 度 为 0.02% 的叠氮钠(N a N 3),小量分装,置-20℃冻存或冷冻干燥保存。
3 ) 注意事项
(1) 免疫用的动物多为哺乳类和禽类。哺乳类■主要有家兔、豚鼠、猪及猴等。禽类常用鸡。
(2) 免疫动物所用的抗原要求尽量纯化。
( 3 ) 目前用得最多的是弗氏佐剂,而弗氏佐剂又分为完全型和不完全型。
(4) 免疫动物需根据抗原的免疫原性强弱及抗原用量的多少,制定免疫方案。免疫的方案应包括注射的抗原量、途径、间隔时间和免疫次数。
单 克 隆 抗 体 的 制 备
单克隆抗体(monoclonal antibody, M c A b )有高度的均一性,特异性强。它是由抗原致敏的一个B 淋巴细胞增殖而来的细胞系所产生的抗体。这是因为单抗是来源于 — 个 B淋巴细胞的细胞株,而且其只对一种特定的抗原决定簇起反应,所以它具有高度均一性和特异性。不仅如此,杂交瘤细胞还能在体外或小鼠的腹腔内培养,产生大量抗体,易于标准化。
1 ) 所用设备与试剂
注射器、无菌培养皿、组织研磨器、计数器、恒温水浴、离心机、 2 4 孔板、二氧化碳孵箱、 B A L B /C 小鼠、弗氏完全佐剂、无菌生理盐水、 7 5 % 乙醇、 R P M I -1640培养液、5 % 小 牛 血 清 1640液 、台盼蓝、 5 0 % P E G 溶 液 、 2 0 % 小牛血清H A T 选择培养液。
2 ) 操作步驟
(1)动物免疫:常 选 用 B A L B /C 小 鼠 ,进行免疫,抗原用量根据所制备的抗血清的要求不同而异。常 用 12.5〜50 ug/100ul, 与等量弗氏完全佐剂混合,制成乳剂,用 200ul做动物背部皮内多点注射,如果注射量大,可做皮下注射 。 一 般间隔1——3周注射一次,共 3〜 5 次 ,末次接种后一周,切断小鼠尾巴尖部,放 血 ,检测抗血清的特异性和效价。选择产生高质量抗血清的小鼠作为脾细胞的来源。被选小鼠在试血后的 3〜6天,用同一剂量的抗原以无菌生理盐水溶解做静脉加强注射一次,使抗原刺激的B 淋巴细胞定位于脾脏。 3〜 4 天 后 ,处死动物,制备脾细胞。
⑵ 脾 细 胞 制 备 :将免疫的小鼠处死,浸 人 7 5 % 乙醇, 5m i n 换乙醇一次,在无菌条件下取出脾脏,用 R P M I -1640培养液冲洗表面血液,去包膜,将脾脏放入盛有培养液无菌培养皿中,剪 碎 移 入 15m l 的组织研磨器中,加 新 配 的 5 % 小 牛 血 清 1640液 ,挤压脾组织块,制备细胞悬液,收集细胞悬液静置片刻,待组织碎片下沉后,吸 取 上 含 细胞的悬液用5 % 小 牛 血 清 1640液 洗 2 次 ,沉淀再悬浮于Iml 培养液中并用台盼蓝染色,计 数 ,要求活细胞占9 0 % 以上。
(3)骨髓瘤细胞制备:从处于对数生长期的 N S -I 小鼠骨髓瘤细胞株培养液中取细胞2xl0<sup>7</sup>〜 4xl0<sup>7</sup 个 ,加 5 % 小牛血清1640液离心lOmin,去上清,将沉淀重悬计数。
⑷ 细 胞 融 合 :取 脾 细 胞 IO8和骨髓瘤细胞 2xl0<sup>7</sup——4xl0<sup>7</sup 个 ,放 入 50m l 试管中,加R P M I 164 0 液 离 心 ,去上清,使管底沉淀松散,混匀,置 37 丈水浴中。取 50% P E G溶液0.8ml,边搅拌边加入,加完后继续搅拌1.5——2min,使细胞凝聚。加1640培养液,再加未温浴含5 % 小 牛 血 清 1640培养液稀释,终 止 P E G 作 用 ,然 后 离 心 lOmin,去上清加 2 0 % 小牛血清H A T 选择培养液 (含青霉素和链霉素各l00U /ml) 30m l ,饲 养 细 胞 IOml(含细 胞 数 为 2xl0<sup>7</sup〜 4xl0<sup>7</sup个/m l , 饲养细胞可用小鼠腹腔巨噬细胞或未免疫小鼠脾细胞)混匀 ,分 配 到 2 4 孔 板 ,每孔加〇.5m l ,置 3 7 ℃ 含 5 % 〜 7 % 二氧化碳孵箱中培养。经 H A T 选择性培养后,融合的杂交瘤细胞生长良好, 而亲代的脾细胞和骨髓瘤细胞死亡。培 养 7〜 10天 后 ,在倒置显微镜下观察,见融合细胞已覆盖1/3〜 1/4孔底面积时,开始测抗体活性,应反复测定2〜 3 次 ,以确定抗体阳性孔。选出抗体阳性孔的细胞 , 一 部分做克隆化培养,一部分用液氮冷冻保存。
(5) 克隆化培养:常用有限稀释法 (limiting dilution)将抗体阳性孔的细胞进行准确计数 ,使细胞数达到10 个/m l ,然后根据所做9 6 孔板的数量,计 算 所 需 15% 小 牛 血 清 1640培养液与杂交瘤细胞的量,将细胞悬液分配到9 6 孔 板 ,每 孔 0.1 ml。使平均每孔有1个细 胞 ,再加入饲养细胞,一起 培 养 7〜 14天 ,待细胞克隆出现,并 覆 盖 1/3〜 1/4孔底时,再次检测培养液中的抗体活性。并将抗体阳性克隆化细胞再克隆培养 。 一 般至少应进行再克隆培养 2 次以上,按这种方法分配,将得到由单个细胞产生的克隆。
(6) 扩大培养:当找到抗体阳性克隆,应立即扩大培养,收 获 M c A b 。扩大培养制备 M c A b 的方法有:①抗体阳性细胞体外培养,收集培养液,但产量甚微,②腹水培养阳性细胞。具体操作是将无菌石蜡油0.5m l ,注入小鼠腹腔,造成无菌性^症 ,于 注 射 后 1〜 2 周 ,取 IO<sup>6</sup〜 IO<sup>7</sup个抗体阳性细胞用〇.5m l 生理盐水稀释,注入小鼠腹腔,待产生大量腹水后,用注射器抽取腹水,每2〜3 天抽一次,可得到大量的 M c A b 。再经双向免疫扩散,免疫电泳及聚丙烯酰胺圆盘电泳,进行抗体鉴定,然后冷冻干燥,-20℃保存。
(7) 冷冻保存与复苏:冷冻应采用逐渐降温法,各实验室使用的方法大同小异。将细胞装入冰存管,加用小牛血清配制的1 〇%——15% 二甲亚砜(dimethyl sulfoxide, D M S O ),作为细胞保护剂。再将细胞从室温转到4℃ 、 -20℃ 、 -4〇℃ :,在-4O ℃ 放置6——2h处后直接放入液氮中。复苏时,将盛有细胞的冻存管从液氮中取出,立即投入 3 7 ℃水浴中,见已冻融迅速取出,将 细 胞 转 入 IOml离心管中,加 IOml 完全培养基, 1200r/m i n 离 心 5 min,然后将细胞转人培养瓶中培养。
3 ) 注意事项
(1) 作为免疫的小鼠应与骨髓瘤源自同一品系,这样克隆化的杂交瘤细胞,接种到小鼠才易于增殖,且容易获得高质量的单克隆抗体。
(2)挤压脾组织块,制备细胞悬液时只能挤压不能研磨。
(3) 用 来 制 备 M c A b 的 骨 髓 瘤 细 胞 是 B A L B /C 小 鼠 的浆细胞瘤,瘤细胞内缺乏>H G P R T 酶。 目前已筛选出许多不同的骨髓瘤细胞株。制 备 M cA b 应选择不分泌或不合成 小 鼠 Ig 重链、轻链的骨髓瘤细胞株,常用的有 NS-1、 SP2/0 和 653 等 。
(4) 检 测 抗 体 活 性 ,可用放射免疫法,免疫组织化学法、血凝法和细胞毒试验等手 段 。
###细 胞 转 染
在研究中会需要将外源 D N A 导入不同类型的细胞,从而分析其表达并在细胞水平上研究它对细胞生长的影响。外 源 D N A 通常包括基因组 D N A 、克隆的目的基因和寡核苷酸等。实验中常用的方法主要分为两种:物理化学法和病毒载体法。在物化方法中除D E A E -葡聚糖介导法只适合瞬时转染外,其他各种物化方法既可用于瞬时也可用于稳定转 染 ;而不同的病毒载体法,有的适用于稳定转染 (如逆转录载体法),有的适用于瞬时转染 (如腺病毒载体法)。
实验中比较常用的是磷酸钙沉淀法和 DEAE-葡聚糖介导法、电穿孔法、脂质体介导法。
物 理 化 学 方 法
磷酸钙沉淀法
磷酸钙沉淀法的机制还不完全清楚,可 能 是 Ca<sup>2+</sup> -D N A 结合物形成的颗粒沉积在培养细胞的表面,导致细胞的非特异性内吞作用。
H E P E S 缓冲溶液系统的磷酸钙沉淀法 一 一 用 该 方 法 转 染 的 D N A 既可以是基因组D N A 也可以是质粒 D N A 。
1 ) 所用设备与试剂
C O 2 培养箱、无菌培养皿、离心管、培养基、 CaCl2(2.5mol/L )、 2xHEP E S -NaCl2 溶液 (HBS 组 成 : 280 mmol/L N a C l , 10 mmol/L K C l , 1.5 mmol/L N a 2 H P 〇 4, 12 mmol/L 葡萄
糖 , 50 mmol/L H E P E S )
2 ) 操作步骤
⑴ 转 染 前 2 处 内 ,按 Ix lO <sup>6</sup> 个细胞/培养皿的密度接种 IOOmm 的培养皿, 3 7 ℃、 5 %C O 2 培养过夜。在加入沉淀前 2〜 4 h ,换用新鲜的培养基培养细胞。
⑵ 将 需 要 转 染 的 D N A 用乙醇沉淀 (10——50ug/皿) , 空 气 中晾干。将 D N A 沉淀重悬于 450ul 灭菌水中,加 入 2.5m ol/L C aC l2 50ul。
(3) 取 2x H B S 500ul 于一无菌的锥形管中,一边 吹 打 2x H B S ,一边逐滴加入 D N A /CaCl2 溶液,随即祸旋 5s,将其在室温下静置 20 min 以形成沉淀。
(4) 将沉淀均勻加入长满细胞的 IOOm m 培养皿中,轻轻晃动。
(5) 在 37℃ 、 5 % C 〇 2 培养箱中培养细胞 4〜 16 h 。除去培养基, P B S 洗 细 胞 2 次 加IOml 完全培养基培养细胞。
(6) 对于瞬时转染,可在转染后 48——7211 收集细胞,并进行表达分析。对于稳定转染,让细胞生长 2 个倍増时间 (当被转染细胞是贴壁细胞时,倍增时间一般为 1 8 〜2 4 h ) 后分乂培养皿中进行培养。
3 ) 注意事项
⑴ 转 染 用 D N A 需进行纯化,不纯将影响转染效率。
(2) D N A -磷酸钙的比例不当会影响转染效率。
(3) 转染质粒 D N A 时可加入特定的基因组 D N A (载体 D N A ) 提高转染效率。
DEAE-葡聚糖介导法
二乙胺乙基葡萄糖简称 D E A E -葡聚糖,是在生理 p H 条件下带正电荷的多聚阳离子试 剂 ,它能促进靶细胞摄入外源 D N A ,其机制推测是 D N A 与 D E A E -葡聚糖结合易于被靶细胞内吞,并减少细胞内核酸酶对 D N A 的降解作用。该法只适用于瞬时转染,所需D N A 比磷酸钙沉淀法少,且不使用载体D N A 。
1 ) 主要设备与试剂
C O 2 培养箱; 无菌培养皿; 离心管;Tris 缓冲盐溶液 (T B S )(溶 液 A :NaCl 8 g ,KCl 0.38 g ,Na2 H P O 4 0.2 g , Tris 3 g ,用 H C l 调 节 p H 至 7.5, 定 容 至 100m l ,过滤除菌。溶 液 B :CaCl2 • 2 H 20 1.5 g , MgCl2 • 6 H 20 lg,定容至 100m l ,过滤除菌。取上述 IOmI A 液加入三 蒸 水 89m l ,再 加 Iml B 液体,即配制好 T B S ,使用前过滤除菌)。
2 ) 操作步骤
(1) 取 5xl0<sup>5</sup> 个细胞接种于I O O m m 培养皿, 37℃、 5 % C 0 2 培 养 3 天 ,达 到 30%——50%融合。
(2) 乙醇沉淀 4 ug 质 粒 D N A ,空气干燥,溶 于 40 ul T B S 中。
(3) 吸去细胞培养上清液,用 I x PBS 洗涤细胞,加入完全培养基。
⑷ 边 摇 动 边 将 D N A 缓慢加入 80 ul预热至 37℃的 10 mg/ml D E A E -葡聚糖溶液中,再 将 120ulD N A /D E A E -葡聚糖混合液逐滴加人每个培养皿中,轻轻转动培养皿使其分布均 匀 ,置 3 7 ℃ 、 5 % C 0 2 培 养 4 h 。
(5) 吸 去 含 D N A /D E A E -葡聚糖的细胞培养基,加 人 5m l 含 1 0 % D M S O 的 lxPBS,室 温 静 置 Imin , 吸 去 含 D M S O 的 P B S , 用 5 ml P B S 漂洗细胞一次,加 IOml完全培养 基 继 续 培 养 48——72 h 。根据实 验 目 的 ,在适当的时间收获细胞,用于分析基因的瞬时 表 达 。
3 ) 注意事项
(1) D E A E -葡聚糖有较强的毒性,所以常采用低浓度 (1 〇〇 ——400 ug) 长时间 (4〜8h)的方案。
(2) 为了提高转染效率可以加入氯奎,它 可 以 保 护 D N A /D E A E -葡聚糖不被溶酶体降解。
电 穿孔法
该法是将细胞置于高压电场,在电脉冲的作用下,使细胞膜瞬间出现微小的孔洞,使 外 源 D N A 进入细胞。影响电穿孔法基因转移效率的因素有多个,其中电脉冲影响最大 。电穿孔转染的效率取决于受体细胞的类型、转染用的DNA构 象 等 。
1 ) 主要设备与试剂
低温离心机、电穿孔用的电击仪和电击池、含或不含有选择药物的完全培养基电穿孔缓冲液 [可根据实验所用的细胞类型,选用不同的电穿孔缓冲液,一般无血清和抗生素的细胞培养基,其电击条件为:电 容 960 uF ,电 压 250〜 450V (625——1125V /cm。
2 ) 操作步骤
(1) 培养靶细胞至对数生长中期, 4℃、 680 g 离 心 5m i n 收集细胞。
(2) 将细胞沉淀用预冷的电穿孔缓冲液重悬洗涤丨〜2 次 , 4 ℃ 、 6 8 % 离 心 5 min收集细胞。
⑶ 对 于 稳 定 转 染 ,将 细 胞 以 IxlO<sup>7</sup> 个/m l 的密度重悬于预冷的电穿孔缓冲液中。对于瞬时表达,可用更高密度的细胞。
⑷冰上放置所需数量的电击池,分 0.5m l 细胞悬液/池 。
(5) 将线状或超螺旋的 D N A (D N A 应经过纯化) 加入冰上放置的含细胞悬液的电击池中。握住电击池的两侧,晃动以混匀 D N A /细胞悬液,在冰上放置 5 min。
(6) 将电击池放人电击仪的池架上 (室温),用所设定的电压及电容值电击一次或多次 。电击的次数及电压和电容的设定随细胞类型而异,需进行摸索。
(7) 将电击池置于冰上 lOmin。
(8) 用非选择性完全培养基 2 0 倍稀释被转染细胞,并用该培养基洗电击池数次,以移出所有的被转染细胞。然 后 ,于 3 7 ℃ 、 5 % C O 2 培养细胞。
(9) 瞬时转染可于培养 48——72 h 后收获细胞,以分析基因的表达,稳定转染实验可采用含有特殊药物的选择性培养基进行抗性细胞克隆的筛选。
3 ) 注意事项
(I) D N A 浓度应控制在 1——40 ug/10<sup>7</sup> 个细胞范围内,且 线 状 D N A 转染成功率较高。
⑵目前对于动物细胞进行电穿孔 ,一 般采用长脉冲/低电场方法。
脂 质 体 介 导 法
将需要转移的 D N A 等生物大分子包裹于脂质体,由于脂质体具有磷脂双层结构 (与细胞膜类似),因此可与细胞膜融合,将 D N A 等生物大分子转入耙细胞。单阳离子脂质 (如D O T M A 、 D O T A P 等) 及中性磷脂 (常见的为 D O P E ) 可 与 D N A 形成类脂复合物,从而使D N A 更有效地进人细胞。目前阳离子脂质转染试剂已商品化,转染率比其他转染方法更高 ,重复性更好。
1 ) 使用设备与试剂
培养皿、 C O 2 培养箱、培养基、商品化阳离子脂质转染试剂。
2 ) 操作步骤
(1) 以 1x 10<sup>5</sup> ——3x 10<sup>5</sup> 个/孔 在 6 孔 板 或 35m m 培养皿中接种生长状况良好的靶细胞,于 37℃ 、 5 % C O 2 培养箱中培养 18〜 24 h ,至 细 胞 50%——80% 汇 片 。
(2) 在 E p 管中配制 D N A /脂质体复合物,体系如下:稀释 1——2 ug 质 粒 D N A 至 1(%1无 血 清 D M E M 培养基中。同 时 稀 释 2 —— 2 5ul阳 离 子 脂 质 于100ul 无 血 清 d m E M 培养基中,然后将二者混合,轻轻混匀。在室温下孵育 15〜 45 min,使 D N A 与阳离子脂质结合。
(3) 吸去细胞的原培养基,用 2m l 无血 清 D M E M 培养基洗细胞一次,在每一个孔或35m m 培养皿中加入 0.8m l 无 血 清 D M E M 培养基,然后逐滴加入上述 D N A /脂质体复合物 ,并使之均匀分布。于 3 7 ℃、 5 % C O 2 培养箱中培养 3〜 8 h ,一 般 为 5 h 。
(4) 每孔或直径 35m m 培养皿中加入 Iml 含 2 0 % 胎牛血清的 D M E M 完全培养基,置3 7℃ 、5 % C O 2 培养箱中再培养 18——24 h 。此过程中若有细胞毒性,必须更换新鲜的含 10%胎牛血清的 D M E M 完全培养基。
(5) 吸 去 DNA/脂质体复合物的 DMEM 完全培养基,每孔加人 2m l 新 鲜 的 含 10% 胎牛血清的 DMEM 完全培养基,再 培 养 24〜 48 h。
(6) 瞬时转染实验可在转染开始后 48——72 h 收获细胞,以分析基因的表达;稳定转染实验可于转染后 72 h ,将 细 胞 按 I : 1 0 分入含有特殊药物的选择性完全培养基进行抗性细胞克隆的筛选。
3 ) 注意事项
(1) 一般先加入无血清培养基,再加入未稀释的阳离子脂质体试剂。
(2) 如果待转染的细胞不耐受无血清培养基,可以先在无血清培养基中制备 D N A /脂质体复合物,然后将其加入含血清的完全培养基中进行转染。
病 毒 载 体 法
逆转录病毒载体法
基因组结构简单,易于操作,感染宿主范围广且效率高,但是只能感染分裂复制的细胞,滴度较低。此外,在包装细胞中,逆转录病毒载体有可能通过同源重组未野生型辅助病毒,有潜在的致癌危险性。
1 ) 使用设备与试剂
C O 2 培养箱、培养皿、培养板、克隆环、培养基、 H E P E S 缓冲盐溶液 (H B S ) 其组成如下 [137 mmol/L N a C l , 5 mmol/L K C l , 0.7 mmol/L N a 2 H P 〇 4, 6 mmol/L 葡萄糖, 21 mmol/L
H E P E S , 2mol/LCaCl2, 800 昭/m l (100x) 聚凝胺 (过滤除菌),含 15% 甘油的 H B S ]。
2 ) 操作步骤
⑴ 转 染 前 1 天 ,在 I O O m m 培 养 皿 中 1 : 5 或 I : 10 接种包装细胞培养至 10%〜 20%汇片。
⑵ 将 10 略含有药物抗性基因的逆病毒载体质粒 D N A 加 入 预 先 有 0.5 ml H B S 的试 管 中 ,再加入32 ul 2mol/LCaCl2,轻摇。室温下孵育 45min,直到形成细小的淡蓝色沉淀。
(3) 移去包装细胞的培养基,吸 取 H B S -D N A 沉淀轻加在培养皿中央。在超净工作台内让细胞接触 D N A 2Omin, 期间轻轻转动培养皿以均匀分散 D N A 溶液。加 IOmI 培养基 ,将细胞置于 3 7℃ 、 5 % C 0 2 培 养 4 h 。
(4) 吸尽培养基,轻 轻 加 入 2.5m l 室 温 的 H B S /甘油。将培养皿放回培养箱中孵育1.5〜 3.5 min。快速 弃 除 H B S /甘 油 ,用 IOmI 培养基轻轻洗 2 次 ,力卩 5m l 含有血清的培养基 ,培 养 18——24 h 移出培养基,用 0.45 um 的 滤 器 过 滤 ,获得含有短暂产生的病毒;将其保存于- 7 0 ℃或- 8 0 ℃。
(5) 在转染的细胞中加IOml培养基,培养 2〜3 天。
(6) 在 感 染 前 1 天 ,将包装细胞系 [与第⑴步所用的细胞系的 env 类 型 不 同 ] 按 I : 10至 1 : 2 0 传代。
(7) 移去所要感染的包装细胞系的培养基。加病毒 [从第 (5) 步来] 如下,对于一只I O O m m 的培养皿,稀 释 0.1——1.0m l 病毒液至终体积 3——5m l ,并 加 800ug/m l 的聚凝胺至对 于 60mm的 培养皿 ,稀 释 0.1〜 Iml 病毒液至终体积 2m l ,并 加 800ug/m l 的聚
凝胺至终浓度叫 8ug/ml。 3 7 ℃ 、 5 % C 0 2 培 养 I h 以上。
(8) 在 感 染 后 2——3 天 ,按 I : 10 或 1 : 2 0 将感染的细胞传代,加选择性培养基培养 3天 ,换用新鲜的选择培养基,再培养 4——7 天直至可见到细胞克隆。
(9) 用克隆环挑取分离良好的细胞克隆,每个克隆转移至 2 个 2 4 孔培养板或 6 孔培养板的培养孔中,生 长 至 5 0 % 〜 9 0 % 汇片。
(10) 更 换 1/2 体积的培养基,视包装细胞系不同而培养 1——3天,移出培养基,或对其进行滴度测定,或保存于-70℃ 或-80℃ 。
(11) 继续传代各产病毒细胞克隆,直至其可冻存或直至鉴别出最好的产病毒细胞。当鉴定出一个好的产病毒细胞克隆时,加 10%——15% DMSO 在液氮中冻存。
3 ) 注意事项
⑴ 如 果 感 染 持 续 时 间 超 过 6 h 甚至过夜,应使用更多体积的感染混合液,以防细胞脱水。
(2) 待感染靶细胞的接种数和感染混合液用量随培养皿的底面积而改变。
(3) 用几 种 量 的 病 毒 液 (如 0.1m l 、 0.4m l 、 1.0 ml) 感染几只培养皿,对于小鼠逆转录病毒和禽类逆转录病毒的 E 亚群需添加聚凝胺,对于禽类的其他亚群则不需要聚凝胺 。如果细胞对于聚凝胺不敏感 (大多数包装细胞都如此) 则延长培养时间,不会损害细 胞 。
⑷如果细胞对于聚凝胺不敏感,贝_ 培养基以增加体积。对 于 IOOmm 培养皿加培养基至终体积 10m l ,对于直径 60m m 培养皿加4 ml, 培 养 2〜 3 天。
腺病毒载体法
腺病毒可以感染分裂细胞和未分裂细胞,插 入 的 外 源 D N A 容量大。容易获得较高的滴度,病毒颗粒较稳定,但是其不足也比较明显,外源基因表达是暂时的,而且其体内应用容易引起免疫反应等。
1 ) 使用设备与试剂
C O 2 培养箱、培养板或培养皿、人胚肾 (HEK293) 细胞系、阳离子脂质体转染试剂。
2 ) 操作步骤
⑴ 在 6 孔培养板或直径 35m m 培养皿中接种约 5xl05 个 H E K 2 9 3 细胞, 5 % C O 2培养箱中培养24 h 。
(2) 提取重组病毒质粒 DNA 10|ag, 用 Pac I 酶切使之线性化。
⑶ 按 照 厂 家 说 明 用 lipofectamine(GIBCO 公司) 进行转染 293 细胞。
(4) 7〜 10天后用荧光显微镜进行观察,可以看到多数 293 细胞中有 G F P 的表达。
(5) 收取细胞,在- 7 0 ℃ / 3 7℃ 条件下反复冻融 4 次裂解细胞,离心取含病毒的上清再次 感 染 293 细胞以扩增重组病毒。
(6) 3 天后收集细胞,反复冻融 3 次。
(7) 取 400W 稀释液加至 293 细胞培养瓶中,于 3 7℃ 孵 育 3 h ,换新鲜培养基继续培养 18——24 h ,于荧光显微镜下计数 G F P 阳性细胞数,按以下公式计算病毒滴度 :
病毒滴度= G F P 细胞阳性数 X 病毒上清稀释倍数/〇.4 ml (pfu/ml)
3 ) 注意事项
收集毒液后可以进行纯化,用 CsCl 密度梯度超离心法进行病毒的纯化,将浓缩后的病毒储存液作不同比例的稀释。
###定 位 研 究
定位研究的方法主要有荧光显微术、原位杂交、电子显微术。这里主要介绍通过荧光显微镜进行观察研究的方法,其试验策略是利用免疫学和非免疫学的荧光探针所具有的敏感性和特异性来揭示结构与功能的关系。
内源性 酶 检 测
β-半乳糖苷酶活性检测
炔半乳糖苷酶是经典的组织化学报道基因,其活性可以用多种底物来检测。最常用的 是 X -gal。当 々-gal 水 解 X -gal 分子的糖苷键时,会产生一种可溶性的、无色的吲哚单体 。随后,两个游离的吲哚基团无需酶促催化,即可自发氧化形成二聚体。最终形成的卤化靛青是一种极稳定且不溶于水的蓝色复合物。
1 ) 整块组织 (如胚胎) 的 X-gal 染色
⑴ 使 用 设 备 与 试 剂 :含 有 2m m 〇] y L MgCl2 的 P B S 、 2 % 〜 4 % 多聚甲醛、 X-gal 反应缓冲液 (35 mmol/L 亚铁氰化钾、 35 mmol/L 铁氰化钾、 2 mmol/L MgCl2、 0.02% N P -40、 0.01%
脱氧胆酸钠)、 X -gal 储存液 (40 mg/ml X -gal 溶 于 D M F )
(2) 操作步骤:
a. 将组织在冰上预冷的含有 2 mmol/L MgCl2 的 P B S 中剪碎。
b. 在 2%——4% 多 聚 甲 醛 中 冰 上 固 定 30m i n 到几个小时。
c. 用 P B S 漂洗几遍。
d. 用 X -gal 反应缓冲液稀释 X -gal 储存液,与组织在 3 7 1 孵 育 2——4 h 。
e.在 P B S 漂洗多次,直到漂洗液不再发黄为止。最好过夜漂洗。
f. 在明视野的显微镜下观察。
(3) 注意事项:
a. 固定时间不宜太久。
b. 在 X -gal 反 应 中 加 入 N B T 以替代铁离子,会形成紫色沉淀,这一反应比单用X -gal 反应更快更敏感。
2 ) 组织切片的 X-gal 染色
⑴ 使 用 设 备 与 试 剂 :明胶蔗糖包埋液 (7.5% 明胶、 15% 蔗糖、 0.05% 叠氮钠)、包被
载玻片的明胶溶液 (2 g 明胶, O.lg 硫酸铬钾溶于 200m l 水)、 4 % 多聚甲醛、 Gelvatol(①取
208,209-212
蛋白质的结构/结构域。早期 的 M 13 方法和常规的P C R 方 法 ,由于操作繁琐,意外突变多等缺陷,使定点突变的实验受到很大限制,现在实验中常用的是 Stratagene 公司的 QuikChange Site-directed Mutagenesis Kit0
1 ) 操作步骤
⑴ 准 备 突 变 的 质 粒 从 常 规 中 经 纯 化 试 剂 盒 抽 提 质 粒 。
⑵ 设 计 一 对 互 补 的 、带有目标突变的引物,和模板质粒退火后用 PfuTurbo 聚合酶「循环延伸」。 PfuTurbo 聚合酶在非链置换反应条件下使引物线性延伸,产生环状带缺刻的突变产物。
(3) D/m I 酶切延伸产物,由于原来的模板质粒来源于常规大肠杆菌,是 经 d a m 甲基化修饰的,对 I 敏感而被切碎。
⑷带突变序列的质粒由于没有甲基化而不被切开形成具缺刻的双链 D N A ,将其转化 至 X L -2 Blue 超级感受态细胞中,即可得到突变质粒的克隆。
2 ) 注意事项
(1) 循环延伸是指聚合酶按照模板延伸引物 , 一 圈 后 回 到 引 物 5'端终止,再经过反复加热退火延伸的循环,这个反应区别于滚环扩增,不会形成多个串联拷贝。用的是低次数的循环延伸而非 P C R ,有助于减少无意错配。只需要一次酶切和转化。
(2) D/m I 识别序列为甲基化的 G A T C , G A T C 在几乎各种质粒中都会出现,而且不止一次。
磷 酸 化
蛋白质的磷酸化在细胞生长和分化的调控中扮演着重要的角色。靶蛋白的磷酸化程度由它们的蛋白质激酶和磷酸酶的相对活性决定。而且这两类酶的平衡受到许多因素影响 ,如生长因子、激素、细胞周期等。因此研究蛋白质磷酸化水平对于深入了解其所参与的信号转导过程是非常重要的。蛋白质磷酸化的定量测定常用 32P 来完成 (斯佩科特等2001)。
标记
1 ) 所用设备与试剂
细胞培养瓶、液闪计数器、刮 刷 、低磷酸培养基 (无放射性磷酸盐)、裂解缓冲液[20 mmol/L Tris(p H 7.5)]、 2 mmol/L E D T A 和 0.5% S D S 、 N P -40、 DNase I。
2 ) 操作步骤
(1) 用 60m m 或 I O O m m 培养瓶,在完全培养基中培养细胞至所希望的密度。
⑵用预热的低磷酸培养基 (无放射性磷酸盐) 冲洗 2 次 ,然后加人含 0.5〜ImCi<sup>32</sup> P/ml(ICi = 3.7xl0<sup>10</sup> Bq) 的低磷酸培养基 (50——100umol/L 无机憐酸)。
(3) 培养结束后,回 收 <sup>32</sup> P 标记培养基,用 IOml Aquasol(New England Nuclear) 稀 释 ,取一等份用液闪计数器测定 32P 活性。测定培养基中磷酸浓度,从这两个数据计算标记培养基中 32P 比活性。
⑷ 用 无 二 价 离 子 的 P B S 清 洗 32P 标记的细胞2 次。每 个 I O O m m 培 养 皿 加 Iml 预热的裂解缓冲液在沸水浴中裂解细胞。用一次性细胞刮刷将细胞裂解物刮到一边,然后吸到 1.5m l 离心管,热 水 浴 l 〇 min。
(5) 冷却至室温。
(6) 离心留上清,加 仰 - 4 0 至 2 % ,加 MgCl2至5mmol/L。加无蛋白酶的DNaseⅠ 与RNase 各 0.I m g ,室温放置 20 min
3 ) 注意事项
(1) 标准组织培养基有 l——2 mmol/L 磷酸盐。为 提 高 32P 标记效率,通常将培养基中磷酸盐浓度降至 5 〇 umol/L 或者更低。标记时间决定于胞内 A T P 库与培养 基 中 <sup>32</sup> P 比活性达到平衡的快慢。
(2) 对于长时间培养,为避免类似成纤维细胞这样的快速分裂细胞生长减慢,培养基中无机磷酸浓度应保持在 50〜 100 mmol/L 。
(3) 测定培养基中磷酸浓度时也可用高效液相层析从其他 32P 标记的分子中分离A T P ,直接测胞内 <sup>[Y-32P]</sup> A T P 的比活性。
⑷ 2——4 h 的短脉冲标记可以用来研究瞬时的蛋白质磷酸化。然 而 ,在脉冲标记实验中 ,蛋 白 质 <sup>32</sup> P 标记的暂时提高,可能是磷酸化的提高,也可能因为磷酸转换增强,而蛋白质稱联的磷酸的量并没有改变。分别用含有或不含刺激物的样品,用双向凝胶电泳/W estern 印迹比较未磷酸化蛋白质对磷酸化酸性同工型的比率,就可以区别以上两种可
能性。
检 测 <sup>32</sup> P 标记的蛋白质
放射性磷酸 <sup>32</sup> P 发出高能 P 粒子。因此,可 以 用 X 射线片放射自显影检测在单向或者双向凝胶上分辨出来的<sup>32</sup> P 标记的蛋白质。
1 ) 所用设备与试剂
Whatman 3M M 滤纸、液闪计数器、胶片。
2 ) 操作步骤
(1) 单向或双向凝胶电泳分辨 <sup>32</sup> P 标记的样品。
(2) 在 Whatman 3M M 滤纸上吸干凝胶。用放射性墨水在凝胶边缘标几个点。
(3) 在完全黑暗或安全光下,在 X 射线片曝光夹里,将胶片放在凝胶与增感屏之间。确保胶片与凝胶增感屏的闪烁体面紧密接触。让凝胶与胶片夹在两块增感屏之间。用两块塑料板夹紧曝光夹,于- 7 0℃ 曝光。
(4) 冲 洗 X 射线片,显影。
(5) 依放射性记号,在读片机里将考马斯亮蓝染色胶与放射自显影图排列好,标出靶蛋白,拍照。
(6) 切下含祀蛋白的胶条,用液闪计数器检测磷酸化蛋白的放射性。
3 ) 注意事项
放射自显影光密度分析也可以测定蛋白质带的比活性。但是,基于放射自显影对放射性进行精确的定量,需要合适的曝光时间,而且受到胶片狭窄的线性反应期的限制。如 果 有 P-扫描仪,可以用磷光成像板对<sup>32</sup> P 放射性进行定量测定。
蛋 白 质 相 互 作 用
基因是相对静态的,而基因编码的产物—蛋白质,贝 U 是动态的。具有时空性和可
215
2 ) 实验步骤
(1) 将细胞裂解液与 5 0ul 的 5 0 % 谷胱甘肽琼脂糖球珠悬液和 25 飓 G S T 在 4℃ 颠倒混合 解 育 2 h 。
(2) 在微量离心机上以最大速度在 4 ¾ 离心混合物 2 min。
(3) 将上清转移到新的离心管中。
(4) 设定两个含等量上清及 5 0ul 谷胱甘肽琼脂糖球珠的微量离心管。在一管中加约10 ug 的 G S T 融合探针蛋白。两个反应中加入的探针和对照蛋白质的量应当是等摩尔的。将离心管在 4℃ 颠倒混合孵育 2 h。
(5) 在微量离心机上以最大速度离心样品 2 min。
(6) 在新的微量离心管中收集上清。这样样品可通过步骤 (10) 中 的 S D S -P A G E 进行分析。
(7) 用 Iml 冰冷的裂解液洗涤球珠。在微量离心机上以最大速度离心lmin。弃去上清 。重 复 洗 3 次 。
(8) (可选) 加人 50ul 20 mmol/L 的还原型谷胱甘肽 (在 50 mmol/L Tris • Cl p H 8.0 中) 到球珠中,洗 脱 G S T 融合蛋白及任何与其结合的蛋白质。在微量离心机上以最大速度离心2 min
(9) 将球珠 [来自步骤 (7)] 或洗脱蛋白质 [来自步骤 (8)] 与 等 体 积 S D S -P A G E 上样缓冲液混合。
(10) 将样品煮沸 4 min 并 作 S D S -P A G E 分析。
3 ) 结果判定
检 测 与 G S T 融合蛋白结合的蛋白质的方法取决于细胞裂解液是否被 <sup>35</sup> S 标记以及试
验目的。如果目的是检测与融合蛋白结合的所有<sup>35</sup> S标记的蛋白质,就在干胶仪上将胶抽 干 ,并 用 X 射线片曝光进行放射自显影。如果目的是检测特异性结合的蛋白质,就将蛋 白 质 从 S D S 聚丙烯酰胺凝胶转移至膜上,进行免疫印迹分析。如果目的是确定与融合蛋白结合的蛋白质的大小和丰度,而这些蛋白质是来自非放射性的裂解液,就用考马斯
亮蓝或硝酸银将胶染色。
免 疫 共 沉 淀
免疫共沉淀是以抗原和抗体之间的专一性作用为基础的研究蛋白质间相互作用的经典方法之一。在细胞的非变性条件下被裂解时,完整细胞内存在的许多蛋白质-蛋白质结合被保持下来。如果用蛋白质的抗体免疫沉淀该蛋白质,那么与该蛋白质在体内结合的另一蛋白质可能也沉淀下来。基于与蛋白质的生理性相互作用,蛋白质的免疫沉淀归类为免疫共沉淀。这种方法最常用于测定两种目标蛋白质是否在体内结合,但也可用于确定一种特定蛋白质的新的作用搭档。在保持蛋白质与蛋白质相互作用的条件下,收获和裂解细胞,从细胞提取液中特异性地免疫沉淀目的蛋白质,然后通过聚丙烯酰胺凝胶电泳分离免疫沉淀物。该方法的优点是相互作用的蛋白质都是经过翻译后修饰的,处于天然状态,蛋白质相互作用可以在自然状态下进行,能避免人为的影响,分离到天然状态的相互作用蛋白质 (Kay 2003)
217-221
水 和 大 约 3 0 粒无菌玻璃珠。② 把 5 个平板叠放在一起,紧握平板,猛烈地摇动平板直到菌落重新悬浮。③用无菌移液管从每个平板中吸取 5m l 酵 悬 液 。把细胞悬液集中在 50m l 的无菌锥形瓶中。④继续下面 5 个平板,重复①-③ I连 续 从 3 0 个平板中收获酵母细胞,结果得到总体积为 150m l 的酵母悬液,保 存 在 3 个 50m l 的瓶中。
ii.如果需要, 在每个装有酵母细胞的锥形瓶中加无菌 T E (p H 7.5 域无菌水至4〇 〜45m l ,振荡或翻转瓶中悬浮细胞。
iii. 使用台式离心机,室温、 1000——1500 g 离 心 5 min,弃去上清。
iv. 重 复 ii 和 iii。
v. 重新悬浮压紧的细胞沉淀于1 倍体积的无菌甘油溶液中,合并不同管的内容物 ,彻底混合。
Vi.分别转移 Iml 的细胞混合物于一系列无菌小离心管中, - 7 0℃ 冻存。
c.相互作用蛋白的筛选:
i. 解冻一份转化文库酵母细胞,用 C M (Gal-Raff)-Ura-His-Trp 培 养 基 按 1 : 10稀 释 , 3 0 ℃ 摇动培养酵母细胞 4 h 来诱导文库中 G A L l 启动子的转录。
ii.在适当数量的 100m m C M (Gal-Raff)-Ura-His-Trp-Leudropout 平板上分别培养 IO<sup>6</sup> 个细胞。
iii. 30℃ :培养平板 5 天。
iv.观察平板的生长,出现克隆时对其加以标记。
V. 第 5 天,涂布一^按^天出现的不同克隆分组的主平板 C M (Gal-Raff)-Ura-His_Tip。
vi. 3 0℃ 孵育平板直到菌落出现。
d. 阳性相互作用的确定:弟半乳糖苷酶活性和亮氨酸需求的检测。
e. 测定转录活化:①使用一个平头牙签分别在下面的 4 个平板上复制和主平板上相同的栅格。使用相同的牙签在 4 个平板上划线培养单克隆,尽量在含有 X-gal平板上划制粗的酵母线条,而在亮氨酸缺乏的平板上划细的酵母线条。②平板在 30℃:培 养 3〜 4 天 。
f.解释结果。如果具备下面的条件,菌落和它们包含的质粒被认为第一轮的阳性克 隆 :在 C M (Gal)-Ura-His-Trp 平板上的培养物 X -gal 呈现蓝色;在 C M (Glu)-U m -His-Trp 平板上的培养物 X -gal 分析呈现白色或仅是浅蓝色; 菌落在 C M (GalRaff)-Ura-His-Trp-Leu 平板上生长良好;菌落在 C M (Glu)-Ura-His-Trp-Leu 平板上生长缓慢或根本不能生长。
噬 菌 体 展 示 技 术
噬菌体展示技术是将表达多肽的基因与噬菌体表面蛋白编码的基因融合,以融合蛋白的形式表达在噬菌体表面的一种方法,最常用的表达系统是 M 1 3 噬菌体。将 c D N A 文库插入噬菌体载体进行表达后,所得到的总体为噬菌体展示库。为 了 得到与「诱 饵 」蛋白 相 作 用 的 「猎物」蛋白,可将展示库与固定化的「诱馆」蛋白相互作用,则 「猎 物 」蛋白被吸附下来,被吸附的重组噬菌体可经再感染而扩增,从而得到猎物蛋白的插入基
因 ,目前,噬菌体展示技术可用于鉴定和分析与其他蛋白质相互作用的蛋白质的表面特征 ;分离与特定氨基酸序列结合的新的配体;提高配体与靶结构之间相互作用的亲和力和特异性 (Brymora 2004)。
在微量滴定板上进行亲和筛选的方法,是将标靶蛋白固定在微量滴定板上,噬菌体重组肽库加到板上来选择相互作用的蛋白质。淘选过程可以再重复 2 次 ,以富集可结合的噬菌体减少非特异性结合噬菌体的回收。经 过 3 次的淘选,特异性连接的单克隆可通过 E L I S A 进行检测。连接上的可以在加入辣根过氧化物酶标记的抗噬菌体单克隆抗体及显色底物后呈现颜色。
噬菌体表面呈现库的构建主要包括以下的步骤:
(1) 首先对要呈现的目的基因片段进行随机酶切,并对酶切片段进行回收;
(2) 酶切片段和克隆接头进行连接,并插入噬菌体载体;
(3) 重组的噬菌体转人合适的宿主菌建库;
(4) 转化菌培养扩增后提取重组的噬菌体。
试剂与仪器
(I) 0.2mol/L 碳酸氢盐缓冲液 (Na2 H C O 3), p H 8.5;
⑵ 封 闭 缓 冲 液 :含 有 l%(m/V) BSA 的 PBS;
⑶ 洗 脱 缓 冲 液 :含 有 0.05%(W V )Tween-2 〇的 P B S ;
(4) 重组噬菌体展示库;
(5) 酸 洗 脱 缓 冲 液 : 50 mmol/L 甘氨酸-H C 1, p H 2.0;
( 6 ) 中和缓冲液: 0.2mol/LTris • Cl, pH7.5;
(7) 细菌: D H 5a F 培养过夜;
(8) 2x Y T 培养基以及顶层、底层琼脂:
胰蛋白膝 IOg
酵母膏 IOg
NaCl 5 g
溶 于 I L 的水中,高压灭菌。 4 ℃储 存 2 周 ;
(9) 底层琼脂:高压前加琼脂至终浓度为 1.5% (m/V);
(10) 顶层琼脂:高压前加琼脂至终浓度为0.8% (m /V);
(11) 阴性对照蛋白;
(12) 辣根过氧化物酶标记的抗噬菌体 M1 3 的单克隆抗体;
(13) 显色底物;
(14) 9 6 孔微量滴定板;
(15) 移液管;
(16) 5m l无菌管;
(17) 无菌牙签;
.
(18) 能读取微量滴定板的分光光度计;
(19) DNA 纯化测序设备。
方法
1 ) 微量滴定板包被
⑴ 纯 化 的 目 的 蛋 白 1〜 3 ug加 入 〇.5m l 的 0.2mol/L 的碳酸盐缓冲液 (p H 8.5 冲 混 合9 6 孔板中每孔加 1500,在室温下静置孵育 2 h ,少 数 蛋 白 质 可 以 代 过 夜 。
(2) 每孔加入 2 〇〇ul 的封闭缓冲液室温孵育 3 〇 min。
(3) 轻轻倒掉液体,在纸上轻叩两下,去除残留溶液。
(4) 每孔中加人 200叫洗脱缓冲液,洗 孔 3 次。
2) 选择相互作用的嗔菌体
⑴稀释重组噬菌体展示库,每 150ul 洗脱缓冲液含 1000 个噬菌体,之 后 取 150ul加入到孔中,室温孵育 2 h。
(2) 轻轻倒掉液体,在纸上叩干,去除残留溶液。
(3) 加 人 200|il 洗脱缓冲液,洗 孔 3 次。
3 ) 结合嗔菌体的回收
(1) 加 人 150ul 酸洗脱液,洗脱,在室温放置 lOmin,将 3 个孔的液体收集到一个干净 的 5m l 的塑料管中,每孔含有 1〇 <sup>3</sup> ——1〇 <sup>8</sup> 个噬菌体颗粒。噬菌体也可以用〇.2m o l/ L 乙醇胺 (PH12.0) 洗 漆 ,然后中和。
(2) 加 人 200ul过夜培养的细菌,室温孵育 l〇 min。
(3) 加 人 3 ml 2x Y T 顶层琼脂培养基,混勻,倒 人 100个 15m m 的含有 2x Y T 底层琼脂培养基的培养皿中,在室温放置 5 min 凝 固 ,然后将板放于 3 7 ¾ 孵育过夜。(除了用板扩大噬菌体外,还可在液体培养基中扩大。将感染后的细菌的溶液加入到 5m l 的 2x Y T的肉汤中,转 移 至 50m l 的管中,于 37℃ 摇动过夜。)
⑷ 往 顶 层 琼 脂 上 加 人 5m l 洗脱缓冲液,用于洗提噬菌体颗粒,代 静 置 过 夜 。
(5) 取 5 0 0ul 噬菌体悬液13 200r/m i n 离 心 IOmin, 将上清移入新管中,这样就得到了噬菌体。加 400 ul 20%(m /V)P E G /2.5mol/LNaCl 到 1.6 ml 的上清中,冰浴 15 min,迅速
以 13 200r/m i n 离 心 5 min,再 用 2 0 0 0 洗脱缓冲液重悬。
( 6 ) 重复上述过程,以获得大量的噬菌体。每个循环收集到的噬菌体滴度为 IxlO<sup>11</sup> pfu/ml
( 7 ) 收集到的噬菌体在 PB S 中 以 I : 100 稀释。在无菌的 5m l 的管中加入 200ul 过夜培 养 的 D H 5 〇 iF, 1〜 IOul 稀释后的噬菌体, 3m l 的 2x Y T 顶层琼脂培养,混勻。
(8) 将稀释的噬菌体接种在 2x Y T 底层琼脂培养基上, 3 7 ¾ 孵育过夜。
(9) 用无菌的牙签从噬菌斑挑取噬菌体,加 入 到 含 有 50ul 的过夜培养的 D H 5cxF 及lml2x Y T 培养基中。 37℃ 通风过夜培养生长。
4) ELISA 测定
(1) 9 6 孔板中一个孔用标靶蛋白包被,另一个用阴性对照蛋白包被,见 「 1) 微量滴定板包被」的步骤 (1)〜 (4)。 注意设置足够的孔用于检测「 3) 结合噬菌体的回收」中的步骤 (9) 得到的克隆。
⑵每对成对的孔中 (目标与对照伽入 1〇〇ul洗脱缓冲液和 2 5ul 的 「3) 结合噬菌体的回收」中的步骤(9) 得到的单克隆噬菌体的上清,室温孵育 2 h。
(3) 轻 轻 倒 掉 液 体 ,在纸上叩干,去除残留溶液。加 入 20〇 ul 洗脱缓冲液,洗孔3 次 。
⑷ 每 孔 加 l〇〇 ul 的抗噬菌体单克隆抗体(连有辣根过氧化物酶),以 1 : 5000 的比例稀 释 ,室 温 孵 育 lh。
(5) 轻轻倒 掉 液 体 ,在纸上叩干,去除残留溶液。加 入 200ul 洗 脱 缓 冲 液 ,洗孔3 次 。
(6) 每 孔 加 入 IOOul的显色底物,孵 育 3 〇 min。
(7) 4〇5n m 处测量光密度值。
5 ) 确 定 相 互 作 用 的 噬 菌 体 序 列
(1) 插入到重组噬菌体中的 D N A 纯化及测序。
(2) 将编码序列转化为肽序列。
噬菌体展示可用于:①鉴定和分析与其他蛋白质相互作用的蛋白质的表面特征;②分离与特定氨基酸序列结合的新配体;③提高配体与靶结构之间相互作用的亲和力和特异性。
### 蛋 白 质 结 构
蛋白质有成千上万种,不同的蛋白质具有不同的生物功能,生物功能的多样性是由蛋白质结构多样性所决定的。因此,蛋白质结构的研究占相当重要的地位。
蛋 白 质 一 级 结 构 的 测 定 方 法
蛋白质一级结构的测定有助于研究蛋白质的立体结构和蛋白质的功能,了解物种分子进化及鉴定分子病等。
1 ) 蛋 白 质 样 品 的 纯 化
蛋 白 质 样 品 与 等 体 积 S D S - P A G E 上 样 缓 冲 液 混 合 。将 样 品 煮 沸 4m i n 后进行SDS-PAGE 电泳。
2) 测 定 蛋 白 质 分 子 中 的 多 肽 链 的 数 目
通过测定蛋白质分子中 N 端 和 C 端残基的摩尔数及蛋白质的相对分子质量,可以确定蛋白质分子中肽链的数目,但应注意可能有封闭的末端,如乙酰化的 N 端和酰胺化 C端等。
3 ) 多肽链的分离
在测定一个蛋白质的结构以前,首先必须保证被测蛋白质的纯度,使结果准确可靠。其次要了解它的分子质量和亚基数,按照其亚基数将蛋白质分成几个多肽链。
(1) 肽链的拆开。蛋白质分子多肽链的连接有共价结合和非共价结合两种。要拆开以共价结合的一 S- S—连接的多肽链,必须采用的化学处理方法有:
a.过甲酸氧化:用氧化剂过甲酸断裂一 S—S— 。这个反应一般在〇丈下进行 2 h左 右 , 2 个 S 就全部能转变成磺酸基,这样被氧化的半胱氨酸称为磺基丙氨酸。
如果蛋白质分子中同时存在半胱胺酸,那么也会被氧化成磺基丙氨酸。此外甲硫氨酸和色氨酸也可被氧化,从而增加分析的复杂性。
b. 巯基乙醇还原:利用还原剂巯基乙醇亦可使蛋白质的一 S__s 一断裂。当高浓度的巯基乙醇在 P H 8.0 条件下室温保温几小时后,可以使一 s_ s 一定量还原。与此同时反应系统中还需要有 8mol/L 脲 或 6mol/L 盐酸胍使蛋白质变性,多肽链松散成为无规则的构型,此时还原剂就可作用于一 S—S— 。此反应是可逆的,
因此要使反应完全,巯基乙醇的浓度必须在 0.1——0.5moi/L。
c. Cldand 试剂的还原作用 Cleland 指出二硫赤藓糖醇 (dithioerythritol) 及二硫苏糖醇 (dithiothreitol)在氧化还原能力上是比较强的试剂,只 要 0.01mol /L 就能使蛋白质的一 S- S—还原,反应基本与巯基乙醇相似,且在许多球蛋白反应中,可以不用变性剂。 Cldand 试剂首先与蛋白质一 S- S—形成中间物,反应终了,还原剂被氧化形成一个稳定的六环化合物,蛋白质则被还原。蛋白质分子的几条肽链若以非共价键结合,则用尿素、盐酸胍等变性剂即可拆开。
(2) 蛋白质的多肽链被拆开后,将它分离纯化,一般多用凝胶过滤、离子交换、电泳等方法。
4 ) 测 定 多 肽 链 的 氨 基 酸 组 成
(1) 代 测 蛋 白 质 与 5.7 mmol/L 的盐酸在水解管中混合。并加入苯酚以防半胱氨酸和甲硫氨酸氧化,抽去空气,充满氮气, 110℃ 水 解 24 h ,或 在 150℃ 下快速水解 90 min。
(2) 水解得到的氨基酸与化学试剂反应,产生易被检测的氨基酸衍生物。氨基酸与茚三酮反应生成紫色化合物,在 570n m 处有吸收峰。脯氨酸和羟脯氨酸与茚三酮反应生成黄色化合物。
(3) 分离及检测。酸性条件下氨基酸带正电荷,是阳离子。在通过阳离子交换树脂时被吸附到柱上,再用洗脱液洗脱、收集。
5) N 端 和 C 端 氨 基 酸 残 基 的 测 定
N 端分析:①多肽的 a-氨基与异硫氰酸苯酯 (PITC) 试剂耦联;②在无水条件下用三氟乙酸酸裂解,形 成 A T Z -氨基酸;③ A T Z -氨基酸在三氟乙酸水溶液(25%) 中转化成稳定的 P T H -氨基酸;④通过薄层层析、气相色谱、高效液相谱及质谱进行氨基酸的鉴定。 C端分析: C 端自由羧基与乙酸酐反应生成混合酸酐,并进一步环化成曝唑酮, C 端® 唑酮在硫氰四丁胺的作用下,转化为乙内酰硫脲 (T H )。
(1) N 端测定:
a. 二硝基氟苯法(F D N B 、 D N F B ): D N P -氨基酸用有机溶剂抽提后,通过层析位置可鉴定它是何种氨基酸。 Sanger 用此方法测定了胰岛素的 N 端分别为甘氨酸及苯丙氨酸。
b. 氰酸盐法。
c. 二甲基氨基萘磺酰氯法。
(2) C 端分析:
a. 肼解法:这 是 测 定 C 端最常用的方法。将多肽溶于无水肼中, 1 0 0 ℃下进行反应 ,结果竣基末端氨基酸以游离氨基酸状释放,而其余肽链部分与肼生成氨基酸肼。这样羧基末端氨基酸可以采用抽提或离子交换层析的方法将其分出而进行分析。如果竣基末端氨基酸侧链是带有酰胺如天冬酰胺和谷氨酰胺,则肼解时不能产生游离的羧基末端氨基酸。此外肼解时注意避免任何少量的水解,以免释出的氨基酸混淆末端分析。
b. 羧肽酶水解法:羧肽酶可以专一性水解羧基末端氨基酸。根据酶解的专一性不同,可区分为羧肽酶 A 、 B 和 C 。应用羧肽酶测定末端时,需要事先进行酶的动力学实验,以便选择合适的酶浓度及反应时间,使释放出的氨基酸主要是 C端氨基酸。
6 ) 肽 段 的 氨 基 酸 序 列 分 析
E d m a n 降解的最大优越性是在水解除去末端标记的氨基酸残基时,不会破坏余下的多肽链。
⑴ 用 苯 异 硫 氰 酸 酯 (PITC)在 p H 9.0 的碱性条件下对蛋白质或多肽进行处理, PITC与肽链的N 端的氨基酸残基反应,形成苯氨基硫甲酰 (P T C ) 衍生物,即 F T C 肽 。
(2) P T C -肽用三氟乙酸处理, N 端氨基酸残基肽键被有选择地切断,释放出该氨基酸残基的噻唑啉酮苯胺衍生物。
⑶将该衍生物用有机溶剂(如氯丁烷) 从反应液中萃取出来,而去掉了一个 N 端氨基
酸残基的肽仍留在溶液中。萃取出来的噻唑啉酮苯胺衍生物不稳定,经酸作用,再进一步环化,形成一个稳定的苯乙内酰硫脲 (P T H ) 衍生物,即 P T H -氨基酸。留在溶液中的减少了一个氨基酸残基的肽再重复进行上述反应过程,整个测序过程现在都是通过测序仪自动进行。每一循环都获得一个 P T H -氨基酸,经 H P L C 可以鉴定出是哪一种氨基酸。当蛋白质中含有一个或多个半胱氨酸残基时,有时一对半胱氨酸残基会通过二硫键交联。在这种情况下测序,首先要经过处理 (如用过甲酸处理) 切断二硫键,然 后 再 进 行 Edman降解测。
7 ) 重 叠 法 确 定 肽 链 的 一 级 结 构
8 ) 蛋 白 质 分 子 中 二 硫 键 及 酰 胺 键 的 确 定 和 磷 酸
化 、糖 基 化 修 饰 点 的 定 位
(1) 蛋白质分子中二硫键的定位。
a. 蛋白质中由二硫键相连的肽链拆分:
i. 过甲酸处理:蛋白质中带游离巯基的半胱氨酸残基及由二硫键连接的半胱氨酸残基可被过甲酸氧化成黄基丙氨酸残基,将肽链拆分。
ii. 疏基乙醇或二巯基苏糖醇处理:用巯基乙醇或二巯基苏糖醇还原二硫键,再用碘乙酸烷基化,然后分别测定拆分得到的多肽链的一级结构。
b. 酶水解肽链未拆分的蛋白质,找出其中含有二硫键的肽段:蛋白质在微酸环境中进行 (P H 2.0〜 6.5)。 将肽段样品点样在正方形滤纸一角,第一方向碘用分离后,用过甲酸蒸气处理滤纸,然后滤纸旋转 90°,在与第一方向电泳的条件下进行第二方向电泳。由于两次电泳条件相同,不含二硫键的肽段在两次电泳中迁移率相同,最终聚集在正方形滤纸的对角线上。含有二硫键的肽段,在过甲酸处理后产生两个负电性增强的肽段,第二次电泳迁移率与第一次不同,不聚集在对角线上,据此可以加以区分。
(2) 酰胺基的定位。蛋白质的氨基酸组成分析可测定蛋白质中酰胺氮总量。
(3) 磷酸化修饰定位。用 ESI 质谱法或 MALDI 质谱法比较憐酸化前、后肽段的质谱可以定位磷酸化的位点。
(4) 糖基化位点的确定。肽段中糖计划修饰有两种类型,N-糖基化发生在门冬酰胺的侧链 N 原子上, O -糖基化发生在丝、苏氨酸侧链羟基的〇原子上,比较用糖苷酶水解前 、后糖肽的M A L D I M 谱便可以确定修饰位点 (鲁 子 贤 1982)。
蛋 白 质 二 级 结 构 的 测 定
1 ) 圆二色谱分析
圆二色谱分析是检测蛋白质二级结构信息的一种有用的方法。不同的二级结构对应与左旋光和右旋光的光吸收差值。通过分析圆二色谱曲线,可以估计二级结构螺旋、折叠以及无规则卷曲的比值。
样品的处理: Res0urseQ 和 R esourseS 离子交换柱分离的样品,低温中速离心浓缩至0.2 mg/ml,并将缓冲液换至测量缓冲液(浓度为 5 〇 m mol/LTris、 50 mmol/LNaCl、 5 mmol/LMgCl2、 p H 7.5 的缓冲液中含 0. 2 m g/mi) 中,直接用于测量。测 量 杯 长 度 为 0.2 cm , 溶液体 积 l 〇〇 ul。波长频率范围是 190〜 250nm。测量温度为 2 5 ℃
2 ) 动态光散射分析
动态光散射仪 (dynamic light scattering) 分 析 ,在 Protein Solution 公司的 Dyna Pro 仪器上运行。
样品处理: ResourseQ 和 ResourseS 离子交换柱分离的样品,低温中速离心浓缩至0.2 mg/ml,并将缓冲液换至测量缓冲液(浓 度 为 50 mm〇l/L Tris、 50 mmol/L NaCl、 5 mmol/LMgCl2、 pH7.5 的缓冲液中含〇.2 mg/ml 样品)中,取 2〇ul 蛋白质样品, 12 000r/min 高速
离 心 IOmin 后 ,用于测量。在 2 0 次的测量后,计算出溶液状态中的蛋白质的分子半径、分子质量以及主要的颗粒。测量温度为 25 丈 (Wei 1996)。
蛋 白 质 晶 体 空 间 结 构
X 射线衍射法分析蛋白质的晶体空间结构。
1 ) 材料与仪器
R I G U K U 转靶阳极 X 射线发生器安低温液氮控制系统、衍射数据收集的面探测仪、X 射线单色器。
2 ) 方法
X 射线晶体学研究通常采用的 X 射线波长与化学键键长相当,也与晶体内的原子间距离响应,约 为 O.lnm。一个晶体包括上亿个有序排列的基本单元。在晶胞中,原子核外电子组成的电子云对 X 射线产生衍射,由于晶体的周期性衍射束只在特定方向出现,电子云可以用傅里叶级数展开,其中每一项的大小与衍射强度相对应,可直接测量,而每一项的相位目前尚没有成熟的方法直接进行测量(牛 立 文 1998)。
(1) 结 晶 :需要通过大量的条件筛选和优化以恰好是蛋白质分子间的相互作用促成蛋白质分子形成高度有序的晶体,而不是随机聚合形成沉淀。这就要求溶液中的蛋白质处于合适的过饱和状态,并只形成少数的成核中心,使晶体能持续地慢慢生长成大单晶。
(2) 数据收集:通常利用 (单波长)X 射线光束照射在一定角度范围内旋转的蛋白质晶 体 ,同时记录晶体对 X 射线散射的强度。这些强度可转换为结构测定中的结构因子的振幅。
(3)相 角 的 测 定 :结构因子的振幅及相角不是直接相关的量。由于结构因子相角的全部信息在收集数据时丢失,因此必须通过其他途径来得到它们的数值。
(4)相角的改进 (优化):电子密度图的质量及其后的可解释性主要取决于相角的准确性 。有的情况下采用晶胞中不对称单元中的等同部分的电子密度平均,有可能大大地改善误差较大的起始相角。
(5) 电子密度图的解释:相位确定后,可开始计算电子密度图。若从电子密度图能跟踪出肽链走向和分辨出二级结构,则可能推出多肽链的三维折叠方式。进而根据氨基酸序列,就可能构建出原子坐标形式的蛋白质结构模型。
(6) 修 正 :考虑到以建立的立体化学资料的限制,根 据 X 衍射数据对初始的蛋白质分子模型进行修正 (Kensal 1998)。
来源:丁香实验