酵母单杂交技术
互联网
10126
酵母单杂交技术
1.酵母单杂交的基本原理
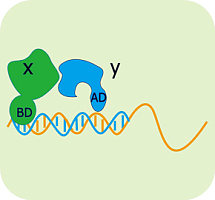
酵母 单杂交技术是1993年由酵母双杂交技术发展而来的,其基本原理为:真核生物基因的转录起始需转录因子参与,转录因子通常由一个DNA特异性结合功能域和一个或多个其他调控蛋白相互作用的激活功能域组成,即DNA结合结构域(DNA―bindingdomain,BD)和转录激活结构域(activationdomain,AD)。用于酵母单杂交系统的酵母GAL4蛋白是一种典型的转录因子,GAL4的DNA结合结构域靠近羧基端,含有几个锌指结构,可激活酵母半乳糖苷酶的上游激活位点(UAS),而转录激活结构域可与RNA聚合酶或转录因子TFIID相互作用,提高RNA聚合酶的活性。在这一过程中,DNA结合结构域和转录激活结构域可完全独立地发挥作用。据此,我们可将GAL4的DNA结合结构域置换为文库蛋白编码基因,只要其表达的蛋白能与目的基因相互作用,同样可通过转录激活结构域激活RNA聚合酶,启动下游报告基因的转录。
酵母单杂交原理示意图
2.酵母单杂交技术的特点
酵母单杂交体系自1993年由Wang和Reed创立以来,在生物学研究领域中已经显示出巨大的威力。应用酵母单杂交体系已经验证了许多已知的DNA与蛋白质之间的相互作用,同时发现了新的DNA与蛋白质的相互作用,并由此找到了多种新的转录因子。近来,已有应用酵母单杂交体系进行疾病诊断的研究报道。随着酵母单杂交体系的不断发展和完善,它在科研、医疗等方面的应用将会越来越广泛。采用酵母单杂交体系能在一个简单实验过程中,识别与DNA特异结合的蛋白质,同时可直接从基因文库中找到编码蛋白的DNA序列,而无需分离纯化蛋白,实验简单易行。由于酵母单杂交体系检测到的与DNA结合的蛋白质是处于自然构象,克服了体外研究时蛋白质通常处于非自然构象的缺点,因而具有很高的灵敏性。目前,多种酵母单杂交体系的试剂盒和相应的cDNA文库已经商品化,为酵母单杂交体系的使用提供了有利的条件。
但酵母单杂交也存在以下缺点:有时由于插入的靶元件与酵母内源转录激活因子可能发生相互作用,或插入的靶元件不需要转录激活因子就可以激活报告基因的转录,因此往往产生假阳性结果。如果酵母表达的AD融合蛋白对细胞有毒性,或融合蛋白在宿主细胞内不能稳定地表达,或融合蛋白发生错误折叠,或者不能定位于酵母细胞核内,以及融合的Gal4AD封闭了蛋白质上与DNA相互作用的位点,则都可能干扰AD融合蛋白结合于靶元件的能力,从而产生假阴性结果。
3.酵母单杂交的基本操作过程
(1)设计含目的基因(称为诱饵)和下游报告基因的质粒,并将其转入酵母细胞。
(2)将文库蛋白的编码基因片段与GAL4转录激活域融合表达的cDNA文库质粒转化人同一酵母中。
(3)若文库蛋白与目的基因相互作用,可通过报告基因的表达将文库蛋白的编码基因筛选出来。在这里作为诱饵的目的基因就是启动子DNA片段,文库基因所编码的蛋白就是启动子基因结合蛋白。
4.酵母单杂交技术的用途
迄今为止,应用酵母单杂交体系已经识别并验证了许多与目的DNA序列结合的蛋白质,同时单杂交技术还被应用于识别金属反应结合因子。正向与反向单杂交体系的结合,还可用于筛选阻碍DNA与蛋白质相互作用的突变的单个核苷酸。
目前,在研究DNA―蛋白质相互作用中,酵母单杂交体系主要有以下3种用途:①确定已知DNA―蛋白质之间是否存在相互作用;②分离结合于目的顺式调控元件或其他短DNA结合位点蛋白的新基因;③定位已经证实的具有相互作用的DNA结合蛋白的DNA结合结构域,以及准确定位与DNA结合的核苷酸序列。