2020-09-21
相关产品推荐
胶原酶 NB 8 来源于溶组织梭菌,9001-12-1,EnzymoPure™, 广谱级,≥ 0.90 U/mg,阿拉丁
¥1544.90
EZElisa™腺病毒-免疫球蛋白A(ADV-IgA)试剂盒-EZElisa™ adenovirus-Immunoglobulin A (ADV-IgA) Kit
¥6448
蛋白质组学 重庆医科大学
¥600
[精选]SCI论文写作投稿资料包:100+资料& 投稿答疑30问
¥0.01
anti-HIV1 p55+p24+p17
¥1180
相关问答
问
HIV胶体金试纸研发问题
求助SCI返修——SCI提交修改稿后发现忘记提交表格
关于最新研究“铜死亡”
相关方法
做 EMSA 时一直发现堵孔的情况,怎么解决?
2023-06-03
基于 FRET 的 HIV-1 病毒粒子的融合试验
2022-02-11
含 BlaM-Vpr HIV-1 病毒粒子的制备
推荐阅读
Sci Adv|半人工光合作用领域取得新突破
Sci Adv:红得发紫的 cGAS,再添新功能!哈佛兰利团队发现它是 DNA 复制减速器
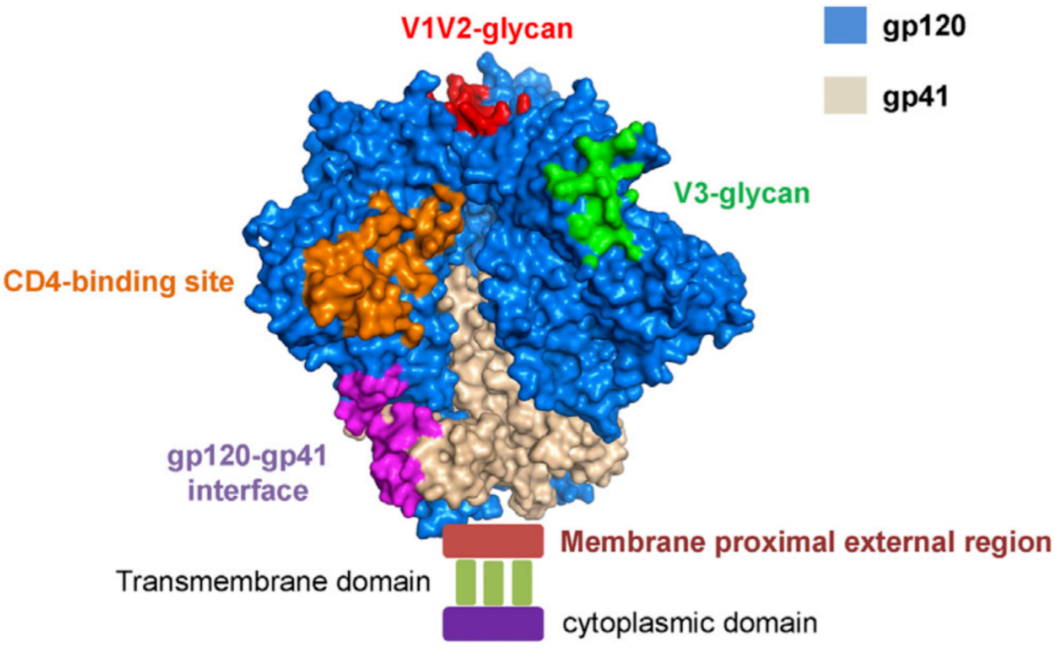
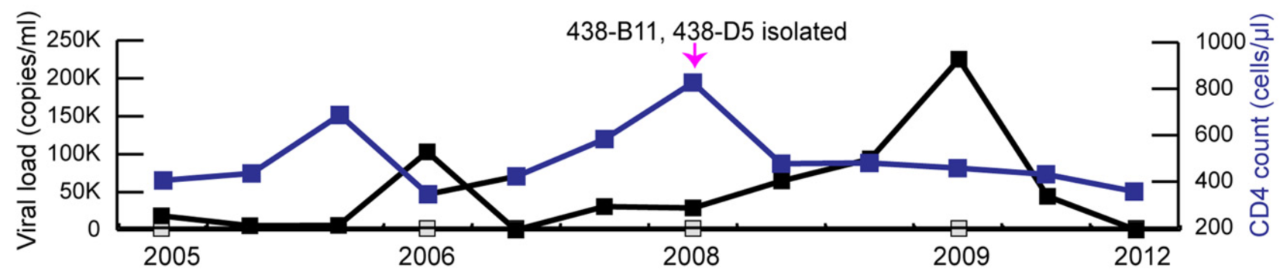
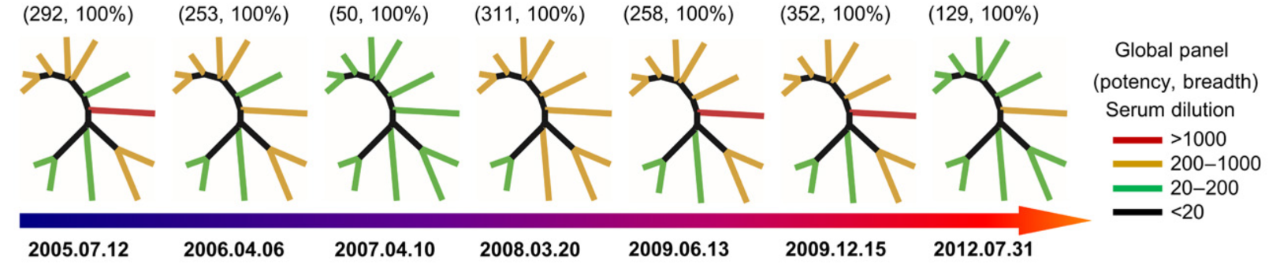
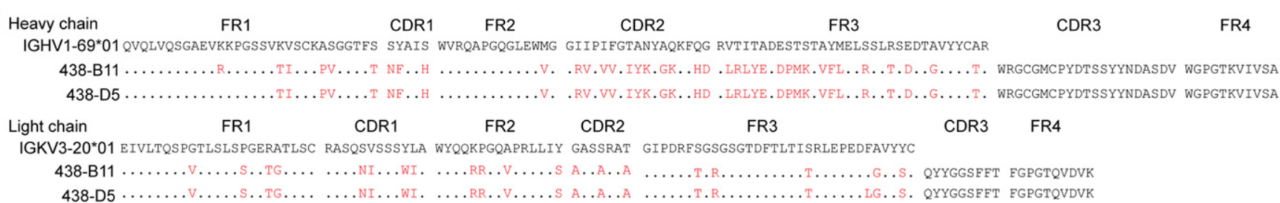
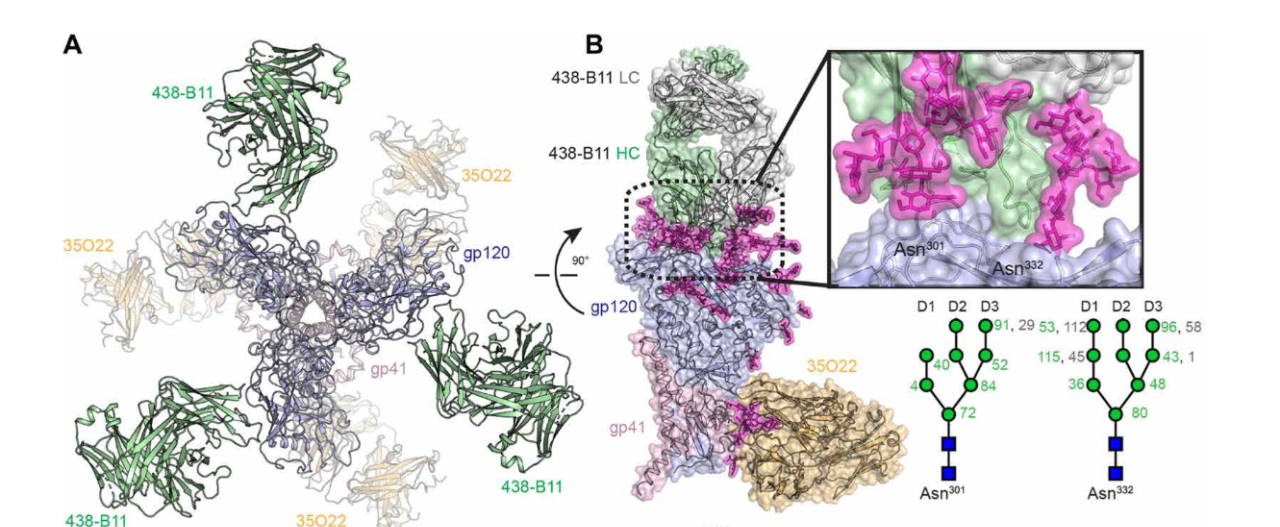
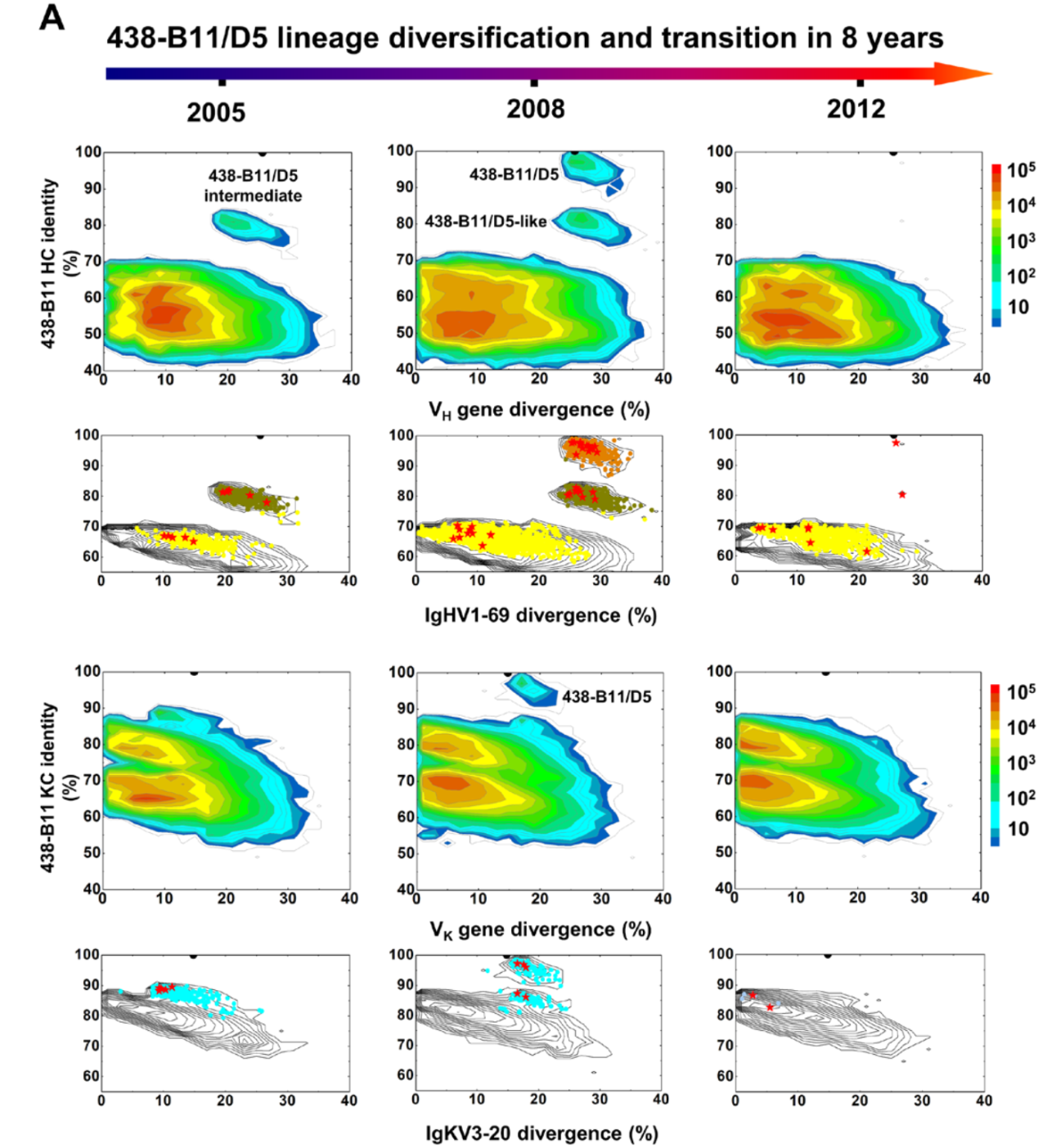
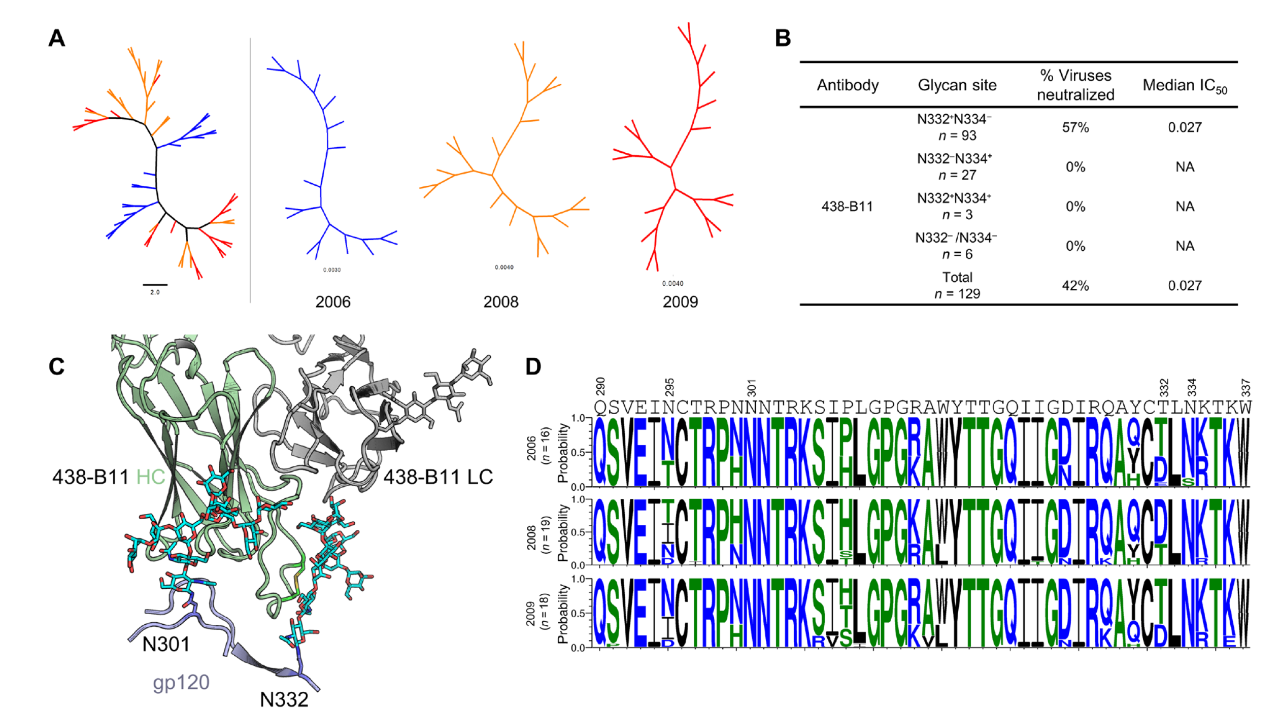
重大进展!Science 同期两篇论文揭示 HIV-1 广谱性中和抗体的产生机制








![[精选]SCI论文写作投稿资料包:100+资料& 投稿答疑30问](https://img1.dxycdn.com/p/s14/2023/1105/569/6102685706544601271.jpg!wh200)



