简介
单磷酸标记可以同时对多个激酶标记,也能在复杂的细胞群体中,标记完成利用细胞表面分子区分不同细胞亚群,同时也检测其胞内磷酸化蛋白。
材料与仪器
步骤
单磷酸标记的基本过程可分为如下几步:
(一)细胞准备
A 细胞培养。按 1×106。细胞/制的细胞量接种于标准组织培养板(6 孔、12 孔和 24 孔)。细胞株常用血清饱和 12 h(时间可不同)。如 Jurkat 细胞尽管是 T 细胞研究的常用模型,但 PTEN 和 SHIP-1 基因缺失导致信号分子主要通过 AKT 和 PI3 激酶效应途径。这种突变使 Jurkat 细胞与初始 T 细胞相比,能在培养时发生细胞增殖,而后者只能在接受外源性刺激后才能增殖。通常, 细胞在高浓度非天然血清中(如牛血清)生长时需要高浓度的生长因子,以提供细胞不能从其天然环境中获得的刺激信号。这样, 它们适应了在非天然环境中生长。在这样的环境中,它们常需要高刺激信号。在这样的条件下,许多信号系统并不反映其基础状态,而背景的活化则变得很明显。在抗体初始滴定时,常用 1 X106 细胞。正确理解某个细胞数的抗体滴度是获得优选信号/噪声比的重要先决条件。其原因是,最佳的抗体浓度和在细胞内结合靶抗原的效力,是反映每个细胞内被结合靶磷酸表位数目、细胞数、非特异性结合阻断剂浓度和能发生背景结合的功能之一。一旦最适条件确定后,细胞数和抗体量也就相应地被确定。如对同样的样本进行多个不同的标记,就有必要对细胞数进行相应放大,以便样本在固定和穿孔后,能被分成几个检测管进行不同染色与处理。
B 加入合适浓度的细胞刺激。除了不刺激/刺激样本外,还要准备对照染色(如同型对照或间标时单独的二抗对照)。再 37℃,15 min。
(二)固定并穿孔
A 直接加入浓缩甲醛溶液于细胞培养物中,使其终浓度达 1%~2%(如用 16% 甲醛溶液,按每样本 l00μL/mL 加入)。漩涡振荡培养板,使细胞均一分布并固定,重新 37℃,15 min。
B 用移液管上下吹打,并使样本细胞完全被移至流式细胞分析管, 冰浴。因其他细胞有固定剂存在,活化细胞更易黏附到塑料上,移液管上下吹打取出大部分这类细胞。通过前后的细胞计数算出丢失的细胞数。通过显微镜观察确定细胞被完全转移,因为培养板中有的地方对于新手来说常会发生明显的细胞丢失。
C 细胞离心(500 g,4℃,5 min)。
D 当细胞以中速漩涡振荡混合时,加入 1 mL 冰冷甲醇到细胞管中,将细胞通透。必须按合理的速率加入甲醇(如 2〜3s/mL)。插入冰中 15 min
E 用 2〜4 mlPBS 洗细胞 3 次,确保将甲醇移去。如甲醇仍然存在,用标记液洗涤常会导致 FCS 蛋白沉淀,要避免这种现象发生。
(三)染色
A 来源于同一刺激条件的刺激细胞需要重新分配以进行不同抗体标记。在标记液中重悬细胞,按每样本 25~100μL 等分细胞。用于抗体标记的细胞浓度应控制在(0.5-1)X106 细胞,并按每样本 25〜100μL 等分。如每标记样本中细胞越少,其在流式细胞仪上的上样时间就越长。因而调整细胞数是很重要的。
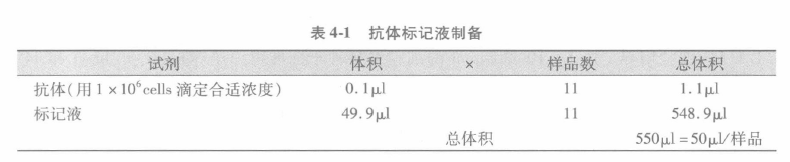
B 加入第一抗体至细胞液,孵育 30 min(有的试剂需要孵育更长时间,如 60 min)。通常我们制备一均质的抗体标记液用于所有的样本。这样,如抗体需要 0.1μL/样本,而我们有 10 个样本,我们就按 N + 1 的样本来制备 50μL 终体积。这样所有的样本都可以用同样体积的试剂来标记。
C 用 1 mL PBS 洗细胞两次后,用 l00 μL PBS-EDTA 重悬细胞。如果是第一抗体直接交联荧光基因,你可以马上在流式细胞仪上对样本进行分析。如果是间接标记(如用了第二抗体),继续 D。
D 按预先测定的稀释浓度加入荧光素标记的第二抗体(根据商品推荐浓度或以 1:500~1000 作为开始浓度),作用 15 min。如你要获得最低的非特异性背景标记,还应对样本做单独的二抗对照。还常检测二抗的滴度,以达到对信号的最大区分或鉴别。
E 用 PBS 洗细胞 2 次后,用 100 μL PBS-EDTA 重悬细胞,在流式细胞仪对样本进行分析。
F 调节流式细胞仪的电压,将同型对照或二抗对照控制在较低水平(如低于 101 log 荧光通道)。然后收获未处理样本,并将处理样本与未处理样本做相对比较。有时, 培养条件会人工活化信号系统(这样需要对大多数细胞株进行血清饱和处理)。如前所述, 这将增加未诱导细胞中一些激酶的水平。
(四)同时对多个激酶标记
如能对细胞内多个活化激酶进行同时检测,将为研究信号分子的级联活化、信号分子间关系提 供更多的机会,并能实时监测其活化状态。我们运用该种方法获得了时间参比试样的相关生物信 号和刺激后动力学变化。另外,如能同时快速检测多个靶信号, 将在高通量药物检测中有明显的优 势, 因为这些方法能检测胞内激酶活性(确保对关键信号组份的药物调节分子跨膜或如预期所想的 作用)和靶标确认(确认对设想蛋白特异性和无其他信号分子对同一条途径中的干扰)。
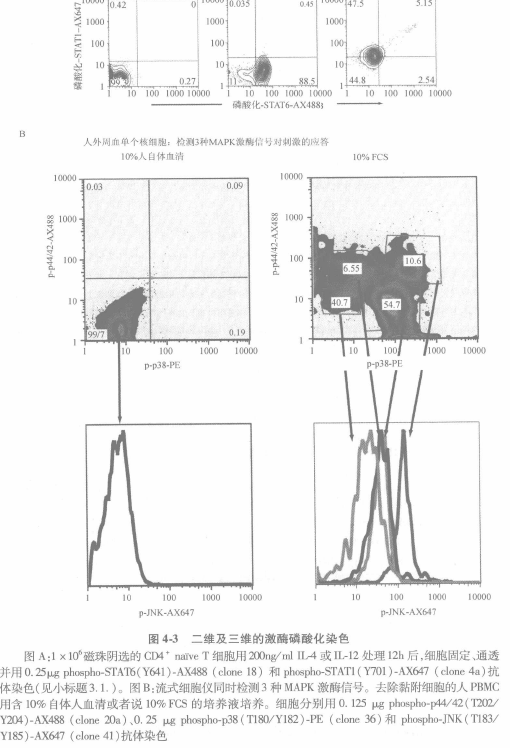
在对缺乏细胞表面分子或其他流式细胞仪标志(如凋亡、DNA 等)分析时,同时对多个磷 酸化蛋白进行分析评估,上述提及的方法已足够。检测用的第二抗体被加入到抗体混合液中(用第一个例子中所列方法进行大致计算),然后将该抗体混合液加到所有的样本中(图 4-3)o 因为多个应用的单克隆或多克隆抗磷酸化抗体使有关染色的考虑因素变得更复杂,所以胞内磷酸化蛋白质检测数量的增加将需要直标抗体进行检测。商品化直标抗磷酸化特异性抗体规 格能满足大多数使用者的要求,然而,某些试剂仍需要实验室自己制备。
(五)表面分子和胞内磷酸标记
在复杂的细胞群体中,我们总希望能运用皂素或甲醇穿孔技术,一步标记法完成利用细胞 表面分子区分不同细胞亚群,同时也检测其胞内磷酸化蛋白。为完成这一目标, 需要对细胞表面分子在固定、穿孔前后的情况做出评估,以确保对表面分子的可检测性。我们在二步标记法中,应用皂素穿孔相关的技术,对免疫分型的细胞亚群内相关磷酸化谱进行了多达 11 个参数的分析。然而,这种方法在应用时,尤其需要注意其中的实验细节,应用磷酸酶抑制剂和冰冷缓冲液控制磷酸酶活性。如用甲醇来完成此类实验,我们发现在某些情况下 需要对表面抗原分子标记进行分析评估。如我们观察到细胞表面抗原表位只能在广泛再水化 后才能被检测时, 就不能应用甲醇来固定了。一些抗原表位因仅松散地保留在细胞上, 极易从 细胞表面剥离。此类抗原表位在应用甲醇固定时,就极易被丢失。如前所述,对于检测表面抗 原表位的抗体还需要进行滴度测定。
通常对于两步标记法,包括连续的胞内和胞外标记两步。对于应用皂素的实验,最好先对表面分子标记,再进行固定与穿孔。在皂素和甲醇的一步标记法中,胞内和胞外抗体最好在固定、穿孔后联合使用。在检测亚群的特异性信号之前,如穿孔后标记会增加着色背景,因而,最好对表面分子的标记差异进行分析与评估(根据所用方法来确定)。
我们完成了一系列表面分子标记实验,其中包括对各种颜色的 200 多种抗体在多聚甲醛、皂素、甲醇固定前后, 穿孔前后的标记能力,及多聚甲醛、皂素、甲醇对荧光素结合蛋白效应和无机染料影响进行了分析。我们的实验结果显示,不能预测何种抗体(克隆或抗原)或荧光素是最好的(0. D. P. 和 G. P. N., 未显示的数据)。多聚甲醛和/或皂素的处理对于大部分颜料没有影响,但甲醇处理对于大部分抗体有影响。例如,图 44 显示,在固定前后和通透条件下对经典的 T 细胞标志 CD4,CD3,CD62L 和 CD8 的检测。固定和皂素通透条件下这些抗体仍保持有抗体活性。固定后标记和皂素通透导致前向散射角减小(细胞大小)。在设门的淋巴细胞内包括有一群荧光参数上调的中间群细胞,通常情况下这群细胞是在淋巴细胞所设门之外的(第四栏)。在此例中,甲醇通透导致了 CD3 染色背景的增加和 CD8 染色的丢失(第五栏)。这样,对于这些单克隆抗体的最好通透/标记条件可在第三栏找到。
来源:丁香实验



![PBS [10X] (Phosphate Buffered Saline) (80mM Na2HPO4, 1.5M NaCl, 20mM KH2PO4, 30mM KCl, pH 7.4)](https://img1.dxycdn.com/p/s14/2025/0305/454/4475752944173517981.jpg!wh200)