一文初解噬菌体
普健生物
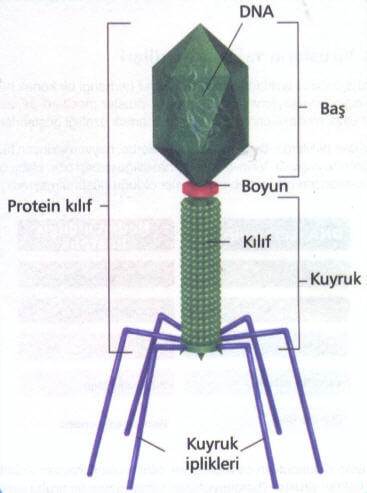
噬菌体是感染细菌、真菌、放线菌或螺旋体等微生物病毒的总称,在电子显微镜下有三种形态:蝌蚪形、微球形和丝形,因部分能引起宿主菌的裂解,故称为噬菌体。跟别的病毒一样,噬菌体只是一团由蛋白质外壳包裹的遗传物质,大部分噬菌体还长有“尾巴”,用来将遗传物质注入宿主体内。
噬菌体颗粒感染一个细菌细胞后可迅速生成几百个子代噬菌体颗粒,每个子代颗粒又可感染细菌细胞,再生成几百个子代噬菌体颗粒。当把细菌涂布在培养基上长成一层菌苔时,一个噬菌体只要感染其中一个细菌,就能把周围的成千上万个细菌感染致死,从而在培养基的菌苔上出现一个因细菌被噬菌体裂解后造成的空斑,称为噬菌斑 (plaque)。
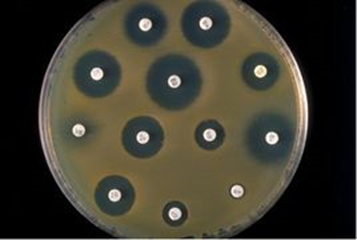
噬菌斑示意图
由噬菌体发展起来的技术也经历了岁月的洗礼:
1985 年,Smith 将 EcoRⅠ的部分基因片段与噬菌体的 fd-tet 基因Ⅲ连接,发现前者编码的多肽能以融合蛋白的形式出现在噬菌体表面,并可与特定的抗体相结合。
1988 年,Parmley 和 Smith 将 B-Gal 表达于噬菌体的表面,并证实该蛋白有生物活性,可以被相应的抗体识别,由此并提出通过构建随机肽库可以了解抗体识别的抗原决定簇表位的设想。
1990 年,Scott 将随机短肽与丝状噬菌体的表面蛋白 PⅢ融合,并展示在噬菌体表面,首次建立了噬菌体随机肽库;McCafferty 用噬菌体展示技术筛选溶酶菌的单链抗体成功使噬菌体展示技术进入一个广泛应用的时代。
随后,许多有功能的蛋白都表达于噬菌体的表面,包括分泌性蛋白(如抗体、生长激素)、细胞内蛋白和酶类,为这些蛋白的基因工程改造奠定了基础。
噬菌体展示系统分类
噬菌体主要有双链噬菌体和单链丝状噬菌体两大类,双链噬菌体为λ类噬菌体,单链丝状噬菌体有 M13、f1、fd 噬菌体。
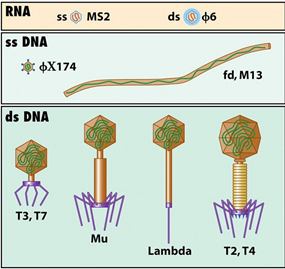
噬菌体分类示意图
接下来主要介绍单链丝状噬菌体展示系统:
PⅢ展示系统
PⅢ是病毒的次要外壳蛋白,位于病毒颗粒的尾端,是噬菌体感染大肠杆菌所必须的。每个病毒颗粒都有 3~5 个拷贝 PⅢ蛋白,其在结构上可分为 N1、N2 和 CT 3 个功能区域,N1 和 N2 与噬菌体吸附大肠杆菌菌毛及穿透细胞膜有关,而 CT 构成噬菌体外壳蛋白结构的一部分,并将整个 PⅢ蛋白的 C 端结构域锚定于噬菌体的一端。当外源的多肽或蛋白质融合于 PⅢ蛋白的信号肽 (SgⅢ) 和 N1 之间时,该系统保留了完整的 PⅢ蛋白,噬菌体仍有感染性;但若外源多肽或蛋白直接与 PⅢ蛋白的 CT 结构域相连,则噬菌体丧失感染性,这时重组噬菌体的感染性由辅助噬菌体表达的完整 PⅢ蛋白来提供。PⅢ蛋白很容易被蛋白水解酶水解,所以有辅助噬菌体超感染时,可以使每个噬菌体平均展示不到一个融合蛋白,即所谓 “单价” 噬菌体。
PⅧ及其他展示系统
PⅧ是丝状噬菌体的主要外壳蛋白,位于噬菌体外侧,C 端与 DNA 结合,N 端伸出噬菌体外,每个病毒颗粒有 2700 个左右 PⅧ拷贝。PⅧ的 N 端附近可融合五肽,但不能融合更长的肽链,因为较大的多肽或蛋白会造成空间障碍,影响噬菌体装配,使其失去感染力。但有辅助噬菌体参与时,可提供野生型 PⅧ蛋白,降低价数,此时可融合多肽甚至抗体片段。 此外,尚有丝状噬菌体 PⅥ展示系统的研究报道。PⅥ蛋白的 C 端暴露于噬菌体表面,可以作为外源蛋白的融合位点,可用于研究外源蛋白 C 端结构区域功能。从所掌握的文献来看,该系统主要用于 cDNA 表面展示文库的构建,并取得了不错的筛选效果。
噬菌体展示文库的用途
1.生产高效低价疫苗
用噬菌体展示某些特异性抗原,可获得低价高效的工程疫苗,而且有研究显示,在 PVⅢ 蛋白的 N 末端展示的多肽,其免疫反应的强度远远高于PⅢ基因蛋白上的外源多肽,而且此法产生的抗体特异性强,因此是生产高效廉价疫苗的好途径。
2. 生产诊断用疫苗
在抗原决定簇已知的情况下,构建重组噬菌体可以为血清学分析提供大量的抗原来源,因为它不需要裂解宿主细胞而直接分泌到培养基中;抗原制备纯化简单,成本较低;噬菌体颗粒稳定,易于长期保存;能在同一个噬菌体同时展示不同的病原体的抗原决定簇。
在预先不知道诊断用靶抗原的情况下,使用患者血清从抗原决定簇库中筛选出展示特异性抗原决定簇的噬菌体,这种血清只能与病人血清抗体反应,而不能与正常人的血清反应,从而建立起疾病的特异性诊断方法。
3.构建 cDNA 文库
一般可用 M13 噬菌体或其他 E.coli 的噬菌体作为 cDNA 文库的展示系统。cDNA 文库的噬菌体展示提供了一个应用免疫学方法进行筛选的条件,取代了细胞内表达 cDNA 文库然后挑选多个菌落的方法。由于 E.coli 只能有效表达一部分真核蛋白,而 M13 噬菌体和其他 E.coli 所能表达的真核蛋白更少。但无论如何,噬菌体表面 cDNA 文库的表达将是研究蛋白质之间相互作用的有用工具。
4.构建噬菌体展示抗体库
抗体库是指免疫球蛋白可变区基因片段的重排而产生的各种抗体的总和。噬菌体抗体库是指在噬菌体表面衣壳蛋白 N 端展示有抗体 Fab(fragment antigen binding) 或 scFv(single chain Fv))。噬菌体抗体展示技术与单克隆抗体杂交瘤技术相比,筛选范围更大(103 VS 107-109), 时间更短(几个月 VS 几周),操作相对简单,无需免疫,费用低,可无限生产,可直接获取基因,应用前景不可估量。更重要的是,它可构建人源抗体库:利用来自人外周血、脾或骨髓淋巴细胞的 cDNA, 用 PCR 的方法扩增出抗体基因,以分泌 scFv 或 Fab 的形式克隆到噬菌体载体中。
5. DNA 结合蛋白研究
噬菌体展示技术可以用于创造一个大的、具有识别不同 DNA 序列的锌指的多肽库。利用这个多肽库,可以研究有关氨基酸序列与 DNA 结合位点之间的识别规则,可以通过设计锌指多肽去控制基因的表达,比如抑制小鼠细胞系中的癌基因,也可以启动表达质粒的基因,或干扰病毒感染周期。
6.信息传递研究
从多肽库中可分离到与天然激素相似的,与受体结合的高亲和力的多肽,因此用完整细胞可以从多肽库中找到受体的高选择性配体。在不知道任何有关受体和配体信息的情况下,用完整细胞和组织或动物,可筛选到特异与靶组织结合的多肽和蛋白。
7.蛋白质组学研究
噬菌体展示技术作为一个多肽和蛋白合成的工具箱,使得任何蛋白均能找到与其有特异结合的多肽和蛋白质,其构建的文库的滴度可达 1012。蛋白质组学的基本点就是利用蛋白质-蛋白质的相互作用,对成千上万的蛋白质同时进行分析,噬菌体展示技术正好能符合这种要求。例如将噬菌体展示 cDNA 文库制成蛋白点阵,成功地应用于寻找与过敏症有关的蛋白质。
噬菌体展示技术优势
(1)淘选的高效率使得在极低的存在水平下挑选高亲和力噬菌体成为可能。
(2)所挑选到的噬菌体可在微量存在的情况下通过感染细菌得到富集。
(3)展示的多肽或蛋白质与包含在噬菌体内部的基因密码的连接,使得结合肽或蛋白质的序列分析既快速又简便。01
受大肠杆菌的转化效率(107-109)的影响,目前常用的噬菌体展示文库中含有不同序列分子的数量一般限制在109。
噬菌体展示技术局限
从建库开始,编码肽的基因就带有一定的偏爱性,从先天决定了肽库多样性的局限性。不是所有的序列都能在噬菌体中获得很好的表达,因为有些蛋白质功能的实现需要折叠、转运、膜插入和络合,导致在体内筛选时需外加选择压力。例如,在噬菌体展示文库试验中,由于部分未折叠的蛋白在细菌中很容易被降解,因此必须小心控制条件,以保证在噬菌体表面展示的文库没有降解。另外,鼠源抗体在噬菌体中表达差,也是体内选择压力的一个例子。真核细胞蛋白在细菌中表达差是因为它们的蛋白质合成与折叠机制不同的缘故。
普健生物可提供各类噬菌体文库淘选技术服务,也可提供噬菌体文库构建及淘选技术服务。