重组DNA的分离、克隆与测序实验手册
互联网
Introduction
This manual is a compilation of many of the everyday methods used in the average molecular biology laboratory, with emphasis on the techniques for large scale DNA sequencing protocols and DNA sequencing automation techniques. The manual has been written in a protocol format, with little theoretical discussion. For theory and additional information, users of this manual are referred back to the original literature, or to other textual manuals such as those published by Maniatis (1) et al. and Glover (2).
The following persons are acknowledged for contributing methods and suggestions during the assembly of this manual: Stephanie Chissoe, Sandy Clifton, Dennis Burian, Rick Wilson, Din-Pow Ma, James Wong, Leslie Johnston-Dow, Elaine Mardis, Zhili Wang, Kala Iyer, Steve Toth, Goughay Zhang, Hua Qin Pan and other members of the Roe laboratory, both past and present.
1. Sambrook, J., Fritsch, E.F., and Maniatis, T., in Molecular Cloning: A Laboratory Manual. Cold Spring Harbor Laboratory Press, NY, Vol. 1, 2, 3 (1989).
2. Glover, D.M. DNA Cloning Volume I: A Practical Approach. IRL Press, Oxford, 1985.
I. General methods
A. Phenol extraction of DNA samples
Phenol extraction is a common technique used to purify a DNA sample (1). Typically, an equal volume of TE-saturated phenol is added to an aqueous DNA sample in a microcentrifuge tube. The mixture is vigorously vortexed, and then centrifuged to enact phase separation. The upper, aqueous layer carefully is removed to a new tube, avoiding the phenol interface and then is subjected to two ether extractions to remove residual phenol. An equal volume of water-saturated ether is added to the tube, the mixture is vortexed, and the tube is centrifuged to allow phase separation. The upper, ether layer is removed and discarded, including phenol droplets at the interface. After this extraction is repeated, the DNA is concentrated by ethanol precipitation.
Protocol
1. Add an equal volume of TE-saturated phenol to the DNA sample contained in a 1.5 ml microcentrifuge tube and vortex for 15-30 seconds.
2. Centrifuge the sample for 5 minutes at room temperature to separate the phases.
3. Remove about 90% of the upper, aqueous layer to a clean tube, carefully avoiding proteins at the aqueous:phenol interface. At this stage the aqueous phase can be extracted a second time with an equal volume of 1:1 TE-saturated phenol:chloroform, centrifuged and removed to a clean tube as above but this additional extraction usually is not necessary if care is taken during the first phenol extraction.
4. Add an equal volume of water-saturated ether, vortex briefly, and centrifuge for 3 minutes at room temperature. Remove and discard the upper, ether layer, taking care to remove phenol droplets at the ether:aqueous interface. Repeat the ether extraction.
5. Ethanol precipitate the DNA by adding 2.5-3 volumes of ethanol-acetate, as discussed below.
B. Concentration of DNA by ethanol precipitation
Typically, 2.5 - 3 volumes of an ethanol/acetate solution is added to the DNA sample in a microcentrifuge tube, which is placed in an ice-water bath for at least 10 minutes. Frequently, this precipitation is performed by incubation at -20C overnight (1). To recover the precipitated DNA, the tube is centrifuged, the supernatant discarded, and the DNA pellet is rinsed with a more dilute ethanol solution. After a second centrifugation, the supernatant again is discarded, and the DNA pellet is dried in a Speedy-Vac.
Protocol
1. Add 2.5-3 volumes of 95% ethanol/0.12 M sodium acetate to the DNA sample contained in a 1.5 ml microcentrifuge tube, invert to mix, and incubate in an ice-water bath for at least 10 minutes. It is possible to place the sample at -20℃ overnight at this stage.
2. Centrifuge at 12,000 rpm in a microcentrifuge (Fisher) for 15 minutes at 4 ℃, decant the supernatant, and drain inverted on a paper towel.
3. Add 80% ethanol (corresponding to about two volume of the original sample), incubate at room temperature for 5-10 minutes and centrifuge again for 5 minutes, and decant and drain the tube, as above.
4. Place the tube in a Savant Speed-Vac and dry the DNA pellet for about 5-10 minutes, or until dry.
5. Always dissolve dried DNA in 10 mM Tris-HCl, pH 7.6-8.0, 0.1 mM EDTA (termed 10:0.1 TE buffer).
6. It is advisable to aliquot the DNA purified in large scale isolations (i.e. 100 ug or more) into several small (0.5 ml) microcentrifuge tubes for frozen storage because repeated freezing and thawing is not advisable.
Notes on precipitation of nucleic acids
A. General rules
Most nucleic acids may be precipitated by addition of monovalent cations and two to three volumes of cold 95% ethanol, followed by incubation at 0 to -70 ℃. The DNA or RNA then may be pelleted by centrifugation at 10 to 13,000 x g. for 15 minutes at 4℃. A subsequent wash with 70% ethanol, followed by brief centrifugation, removes residual salt and moisture.
The general procedure for precipitating DNA and RNA is:
1. Add one-tenth volume of 3M NaOAc, pH 5.undefined to the nucleic acid solution to be precipitated,
2. Add two volumes of cold 95% ethanol,
3. Place at -70℃ for at least 30 minutes, or at -20℃ overnight.
or alternatively
1. Combine 95 ml of 100% ethanol with 4 ml of 3 M NaOAc (pH 4.8) and 1ml of sterile water. Mix by inversion and store at -20℃.
2. Add 2.5 volumes of cold ethanol/acetate solution to the nucleic acid solution to be precipitated.
3. Place at at -70℃ for at least 30 minutes or -20℃ for two hours to overnight.
B. Oligonucleotides
Add one-tenth volume of 3M NaOAc, pH 6.5, and three volumes of cold 95% ethanol.
Place at -70℃ for at least one hour.
C. RNA
Add one-tenth volume of 1M NaOAc, pH 4.5, and 2.5 volumes of cold 95% ethanol.
Precipitate large volumes at -20℃ overnight.
Small volume samples may be precipitated by placing in powdered dry ice or dry ice-ethanol bath for five to 10 minutes.
D. Isobutanol concentration of DNA
DNA samples may be concentrated by extraction with isobutanol. Add slightly more than one volume of isobutanol, vortex vigorously and centrifuge to separate the phases. Discard the isobutanol (upper) phase, and extract once with water-saturated diethyl ether to remove residual isobutanol. The nucleic acid then may be ethanol precipitated as described above.
E. Notes on phenol extraction of nucleic acids
The standard and preferred way to remove proteins from nucleic acid solutions is by extraction with neutralized phenol or phenol/chloroform. Generally, samples are extracted by addition of one-half volume of neutralized (with TE buffer, pH 7.5) phenol to the sample, followed by vigorous mixing for a few seconds to form an emulsion. Following centrifugation for a few minutes, the aqueous (top) phase containing the nucleic acid is recovered and transferred to a clean tube. Residual phenol then is removed by extraction with an equal volume of water-saturated diethyl ether. Following centrifugation to separate the phases, the ether (upper) phase is discarded and the nucleic acid is ethanol precipitated as described above.
A 1:1 mixture of phenol and chloroform also is useful for the removal of protein from nucleic acid samples. Following extraction with phenol/chloroform, the sample should be extracted once with an equal volume of chloroform, and ethanol precipitated as described above.
C. Restriction digestion
Restriction enzyme digestions are performed by incubating double-stranded DNA molecules with an appropriate amount of restriction enzyme, in its respective buffer as recommended by the supplier, and at the optimal temperature for that specific enzyme. The optimal sodium chloride concentration in the reaction varies for different enzymes, and a set of three standard buffers containing three concentrations of sodium chloride are prepared and used when necessary. Typical digestions included a unit of enzyme per microgram of starting DNA, and one enzyme unit usually (depending on the supplier) is defined as the amount of enzyme needed to completely digest one microgram of double-stranded DNA in one hour at the appropriate temperature. These reactions usually are incubated for 1-3 hours, to insure complete digestion, at the optimal temperature for enzyme activity, typically 37℃. See the Appendix for a listing of restriction sites present in the M13 (pUC) MCS and a listing of various restriction enzymes, incubation conditions and cut sites.
Protocol
1. Prepare the reaction for restriction digestion by adding the following reagents in the order listed to a microcentrifuge tube:
sterile ddH2O q.s (where "q.s." means quantity sufficient)
10X assay buffer one-tenth volume
DNA x μl
restriction enzymundefined y μl (1-10 units per ug DNA)
Total volume z μl
Note: The volume of the reaction depends on the amount and size of the DNA being digested. Larger DNAs should be digested in larger total volumes (between 50-100 μl), as should greater amounts of DNA.
Refer to the vendor's catalog for the chart of enzyme activity in a range of salt concentrations to choose the appropriate assay buffer (10X High, 10X Medium, or 10X Low Salt Buffers, or 10X SmaI Buffer for SmaI digestions). Restriction enzymes are purchased from Bethesda Research Laboratories, New England Biolabs, or United States Biochemicals.
2. Gently mix by pipetting and incubate the reaction at the appropriate temperature (typically 37℃) for 1-3 hours.
3. Inactivate the enzyme(s) by heating at 70-100℃ for 10 minutes or by phenol extraction (see the vendor's catalog to determine the degree of heat inactivation for a given enzyme). Prior to use in further protocols such as dephosphorylation or ligation, an aliquot of the digestion should be assayed by agarose gel electrophoresis versus non-digested DNA and a size marker, if necessary.
D. Agarose gel electrophoresis
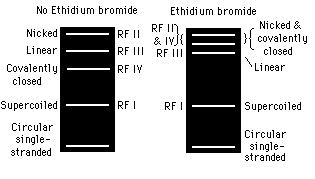
Agarose gel electrophoresis (2) is employed to check the progression of a restriction enzyme digestion, to quickly determine the yield and purity of a DNA isolation or PCR reaction, and to size fractionate DNA molecules, which then could be eluted from the gel. Prior to gel casting, dried agarose is dissolved in buffer by heating and the warm gel solution then is poured into a mold (made by wrapping clear tape around and extending above the edges of an 18 cm X 18 cm glass plate), which is fitted with a well-forming comb. The percentage of agarose in the gel varied. Although 0.7% agarose gels typically are used, in cases where the accurate size fractionation of DNA molecules smaller than 1 kb is required, a 1, 1.5, or 2% agarose gel is prepared, depending on the expected size(s) of the fragment(s). Ethidium bromide is included in the gel matrix to enable fluorescent visualization of the DNA fragments under UV light. Agarose gels are submerged in electrophoresis buffer in a horizontal electrophoresis apparatus. The DNA samples are mixed with gel tracking dye and loaded into the sample wells. Electrophoresis usually is at 150 - 200 mA for 0.5-1 hour at room temperature, depending on the desired separation. When low-melting agarose is used for preparative agarose gels, electrophoresis is at 100-120 mA for 0.5-1 hour, again depending on the desired separation, and a fan is positioned such that the heat generated is rapidly dissipated. Size markers are co-electrophoresed with DNA samples, when appropriate for fragment size determination. Two size markers are used, phi-X 174 cleaved with restriction endonuclease HaeIII to identify fragments between 0.3-2 kb and lambda phage cleaved with restriction endonuclease HindIII to identify fragments between 2-23 kb. After electrophoresis, the gel is placed on a UV light box and a picture of the fluorescent ethidium bromide-stained DNA separation pattern is taken with a Polaroid camera.
Protocol
1. Prepare an agarose gel, according to recipes listed below, by combining the agarose (low gel temperature agarose may also be used) and water in a 500 ml Ehrlenmeyer flask, and heating in a microwave for 2-4 minutes until the agarose is dissolved.
| 0.7% | 1.0% | 2.0% | |
| agarose | 1.05 g | 1.5 g | 3.0 g |
| 20X TAE | 7.5 ml | 7.5 ml | 7.5 ml |
| ddH2O | 142.5 ml | 142.5 ml | 142.5 ml |
| EtBr (5 mg/ml) | 25 μl | 25 μl | 25 μl |
| total vol | 150 ml | 150 ml | 150 ml |
Genetic technology grade (800669) or low gel temperature (800259) agarose from Schwarz/Mann Biotech.
2. Add 20X TAE and ethidium bromide (EtBr), swirl to mix, and pour the gel onto a taped plate with casting combs in place. Allow 20-30 minutes for solidification.
3. Carefully remove the tape and the gel casting combs and place the gel in a horizontal electrophoresis apparatus. Add 1X TAE electrophoresis buffer to the reservoirs until the buffer just covers the agarose gel.
4. Add at least one-tenth volume of 10X agarose gel loading dye to each DNA sample, mix, and load into the wells. Electrophorese the gel at 150-200 mA until the required separation has been achieved, usually 0.5-1 hour (100-120 mA for low gel temperature agarose), and cool the gel during electrophoresis with a fan. Visualize the DNA fragments on a long wave UV light box and photograph with a Polaroid camera.
E. Elution of DNA fragments from agarose
DNA fragments are eluted from low-melting temperature agarose gels using an unpublished procedure first developed by Dr. Roe. Here, the band of interest is excised with a sterile razor blade, placed in a microcentrifuge tube, frozen at -70℃, and then melted. Then, TE-saturated phenol is added to the melted gel slice, and the mixture again is frozen and then thawed. After this second thawing, the tube is centrifuged and the aqueous layer removed to a new tube. Residual phenol is removed with two ether extractions, and the DNA is concentrated by ethanol precipitation.
Protocol
1. Place excised DNA-containing agarose gel slice in a 1.5 ml microcentrifuge tube and freeze at -70℃ for at least 15 minutes, or until frozen. It is possible to pause at this stage in the elution procedure and leave the gel slice frozen at -70℃.
2. Melt the slice by incubating the tube at 65℃.
3. Add one-volume of TE-saturated phenol, vortex for 30 seconds, and freeze the sample at -70℃ for 15 minutes.
4. Thaw the sample, and centrifuge in a microcentrifuge at 12,000 rpm for 5 minutes at room temperature to separate the phases. The aqueous phase then is removed to a clean tube, extracted twice with equal volume ether, ethanol precipitated, and the DNA pellet is rinsed and dried.
F. Kinase end-labeling of DNA
Typical 5'-kinase labeling reactions included the DNA to be labeled, [[gamma]]-32-P-rATP, T4 polynucleotide kinase, and buffer (3). After incubation at 37℃, reactions are heat inactivated by incubation at 80℃. Portions of the reactions are mixed with gel loading dye and loaded into a well of a polyacrylamide gel and electrophoresed. The gel percentage and electrophoresis conditions varied depending on the sizes of the DNA molecules of interest. After electrophoresis, the gel is dried and exposed to x-ray film, as discussed below for radiolabeled DNA sequencing.
Protocol
1. Add the following reagents to a 0.5 ml microcentrifuge tube, in the order listed:
sterile ddH2O .s
10X kinase buffer μl
DNA μl
[[gamma]]-[32-P]-rATP 10 uCi
T4 polynucleotide kinase 1 μl (3U/ul)
10 μl
[[gamma]]-[32-P]-rATP (35020) ICN and T4 polynucleotide kinase (70031) from United States Biochemicals.
2. Incubate at 37℃ for 30-60 minutes.
3. Heat the reaction at 65℃ for 10 minutes to inactivate the kinase.
G. Bacterial cell maintenance
Four strains of E. coli are used in these studies: JM101 for M13 infection and isolation (4), XL1BMRF' (Stratagene) for M13 or pUC-based DNA transformation (5), and ED8767 for cosmid DNA transformation (6-8). To maintain their respective F' episomes necessary for M13 viral infection (9), JM101 is streaked onto a M9 minimal media (modified from that given in reference (1) plate and XL1BMRF' is streaked onto an LB plate (1) containing tetracycline. ED8767 is streaked onto an LB plate. These plates are incubated at 37℃ overnight. For each strain, 3 ml. of appropriate liquid media are inoculated with a smear of several colonies and incubated at 37℃ for 8 hours, and those cultures then are transferred into 50 ml of respective liquid media and further incubated 12-16 hours. Glycerol is added to a final concentration of 20%, and the glycerol stock cultures are distributed in 1.3 ml aliquots and frozen at -70℃ until use (1).
Protocol
1. Streak a culture of the bacterial cell strain onto an agar plate of the respective medium, listed below, and incubate at 37℃ overnight.
E. coli strain Agar Medium/Liquid Media XL1BMRF' (Stratagene) LB-Tet JM101 M9 ED8767 LB
2. Pick several colonies into a 12 X 75 mm Falcon tube containing a 2 ml aliquot of the respective liquid media, and incubate for 8-10 hours at 37℃ with shaking at 250 rpm.
3. Transfer the 2 ml culture into an Ehrlenmeyer flask containing 50 ml of the respective liquid media and further incubate overnight (12-16 hours) at 37℃ with shaking at 250 rpm.
4. Add 12.5 ml of sterile glycerol for a final concentration of 20%, and distribute the culture in 1.3 ml aliquots into 12 X 75 mm Falcon tubes.
5. Store glycerol cell stocks frozen at -70℃ until use.
Notes on Restriction/Modification Bacterial Strains:
1. EcoK (alternate=EcoB)-hsdRMS genes=attack DNA not protected by adenine methylation. (ED8767 is EcoK methylation -). (10)
2. mcrA (modified cytosine restriction), mcrBC, and mrr=methylation requiring systems that attack DNA only when it IS methylated (Ed8767 is mrr+, so methylated adenines will be restricted. Clone can carry methylation activity.) (10)
3. In general, it is best to use a strain lacking Mcr and Mrr systems when cloning genomic DNA from an organism with methylcytosine such as mammals, higher plants , and many prokaryotes.(11)
4. The use of D(mrr-hsd-mcrB) hosts=general methylation tolerance and suitability for clones with N6 methyladenine as well as 5mC (as with bacterial DNAs). (12)
5. XL1-Blue MRF'=D(mcrA)182, D(mcrCB-hsdSMR-mrr)172,endA1, supE44, thi-1, recA, gyrA96, relA1, lac, l-, [F' proAB, lacIqZDM15, Tn10(tetr)].
Host Mutation Descriptions:
ara Inability to utilize arabinose.
deoR Regulatory gene that allows for constitutive synthesis for genes involved in deoxyribose synthesis. Allows for the uptake of large plasmids.
endA DNA specific endonuclease I. Mutation shown to improve yield and quality of DNA from plasmid minipreps.
F' F' episome, male E. coli host. Necessary for M13 infection.
galK Inability to utilize galactose.
galT Inability to utilize galactose.
gyrA Mutation in DNA gyrase. Confers resistance to nalidixic acid.
hfl High frequency of lysogeny. Mutation increases lambda lysogeny by inactivating specific protease.
lacI Repressor protein of lac operon. LacIq is a mutant lacI that overproduces the repressor protein.
lacY Lactose utilization; galactosidase permease (M protein).
lacZ b-D-galactosidase; lactose utilization. Cells with lacZ mutations produce white colonies in the presence of X-gal; wild type produce blue colonies.
lacZdM15 A specific N-terminal deletion which permits the a-complementation segment present on a phagemid or plasmid vector to make functional lacZ protein.
Dlon Deletion of the lon protease. Reduces degradation of b-galactosidase fusion proteins to enhance antibody screening of l libraries.
malA Inability to utilize maltose.
proAB Mutants require proline for growth in minimal media.
recA Gene central to general recombination and DNA repair. Mutation eliminates general recombination and renders bacteria sensitive to UV light.
rec BCD Exonuclease V. Mutation in recB or recC reduces general recombination to a hundredth of its normal level and affects DNA repair.
relA Relaxed phenotype; permits RNA synthesis in the absence of protein synthesis.
rspL 30S ribosomal sub-unit protein S12. Mutation makes cells resistant to streptomycin.
Also written strA.
recJ Exonuclease involved in alternate recombination pathways of E. coli.
strA See rspL.
sbcBC Exonuclease I. Permits general recombination in recBC mutants.
supE Supressor of amber (UAG) mutations. Some phage require a mutation in this gene in order to grow.
supF Supressor of amber (UAG) mutations. Some phage require a mutation in this gene in order to grow.
thi-1 Mutants require vitamin B1(thiamine) for growth on minimal media.
traD36 mutation inactivates conjugal transfer of F' episome.
umuC Component of SOS repair pathway.
uvrC Component of UV excision pathway.
xylA Inability to utilize xylose.
dam DNA adenine methylase/ Mutation blocks methylation of Adenine residues in the recognition
sequence 5'-undefinedATC-3' ~undefined=methylated) dcm DNA cytosine methylase/Mutation blocks methylation of cytosine residues in the recognition sequences 5'-undefinedCAGG-3' or 5'-undefinedCTGG-3' ~undefined=methylated)
hsdM E. coli methylase/ Mutation blocks sequence specific methylation ANundefinedACNNNNNNGTGC or GCNundefinedACNNNNNNGTT ~undefined=methylated). DNA isloated from a HsdM- strain will be restricted by a HsdR+ host.
hsd R17 Restriction negative and modification positive.
(rK-, mK+) Allows cloning of DNA without cleavage by endogenous restriction endonucleases. DNA prepared from hosts with this marker can efficiently transform rK+ E. coli hosts.
hsdS20 Restriction negative and modification negative.
(rB-, mB-) Allows cloning of DNA without cleavage by endogenous restriction endonucleases . DNA prepared from hosts with this marker is unmethylated by the hsdS20 modificationsystem.
mcrA E. coli restriction system/ Mutation prevents McrA restriction of methylated DNA of sequence 5'-undefinedCGG ~undefined=methylated).
mcrCB E. coli restriction system/ Mutation prevents McrCB restriction of methylated DNA of sequence 5'-GundefinedC, 5'-G5undefinedC, or 5'-GNundefinedC ~undefined=methylated).
mrr E. coli restriction system/ Mutation prevents Mrr restriction of methylated DNA of sequence 5'-undefinedAC or 5'-undefinedAG ~undefined=methylated). Mutation also prevents McrF restriction of methylated cytosine sequences.
Other Descriptions:
cmr Chloramphenicol resistance
kanr Kanamycin resistance
tetr Tetracycline resistance
strr Streptomycin resistance
D Indicates a deletion of genes following it.
Tn10 A transposon that normally codes for tetr Tn5 A transposon that normally codes for kanr spi- Refers to red-gam- mutant derivatives of lambda defined by their ability to form plaques on E. coli P2 lysogens.
Commonly used bacterial strains C600 - F-, e14, mcrA, thr-1 supE44, thi-1, leuB6, lacY1, tonA21, l- -for plating lambda (gt10) libraries, grows well in L broth, 2x TY, plate on NZYDT+Mg. -Huynh, Young, and Davis (1985) DNA Cloning, Vol. 1, 56-110.
DH1 - F-, recA1, endA1, gyrA96, thi-1, hsdR17 (rk-, mk+), supE44, relA1, l- -for plasmid transformation, grows well on L broth and plates. -Hanahan (1983) J. Mol. Biol. 166, 557-580.
XL1Blue-MRF' - D(mcrA)182, D(mcrCB-hsdSMR-mrr)172,endA1, supE44, thi-1, recA, gyrA96, relA1, lac, l-, [F'proAB, lac IqZDM15, Tn10 (tetr)] -For plating or glycerol stocks, grow in LB with 20 mg/ml of tetracycline. For transfection, grow in tryptone broth containing 10 mM MgSO4 and 0.2% maltose. (No antibiotic--Mg++ interferes with tetracycline action.) For picking plaques, grow glycerol stock in LB to an O.D. of 0.5 at 600 nm (2.5 hours?). When at 0.5, add MgSO4 to a final concentration of 10 mM.
SURE Cells - Stratagene - e14(mcrA), D(mcrCB- hsdSMR-mrr)171, sbcC, recB, recJ, umuC::Tn5 (kanr), uvrC, supE44, lac, gyrA96, relA1, thi-1, end A1[F'proAB, lacIqDM15, Tn10(tetr)].
An uncharacterized mutation enhances the a - complementation to give a more intense blue color on plates containing X-gal and IPTG.
GM272 - F-, hsdR544 (rk-, mk-), supE44, supF58, lacY1 or