Cell 子刊:颜宁与西湖大学申怀宗团队合作解析 Nav1.7 高分辨率结构,有望助力止痛药研发
丁香学术
在人体中,一共发现了九种电压门控钠离子通道亚型,在不同的器官和生理活动中发挥着不可忽视的作用,钠离子通道异常与神经、肌肉和心血管疾病密切相关,因此解析钠离子门控通道将为药物开发和机制研究提供重要参考。
在九种钠离子通道亚型中,Nav1.7,由 SCN9A 编码,主要在背根神经节神经元中表达,是一种很有希望的止痛药物靶点,在认知信号传导中发挥重要作用。Nav1.7 调节感觉神经元的兴奋性,并有助于几种感官模式的认知。SCN9A 的突变与原发性红热痛、与通道病相关的疼痛不敏感和阵发性极度疼痛障碍有关。
研究人员之前报道过人类 Nav1.7 的结构,在之前的研究中,一种疾病变体 Nav1.7(E406K)由于其增强了重组表达水平而被选择使用。将毒素混合后用于结构解析,最终获得了分辨率均为 3.2 Å 的低温 EM 结构 Nav1.7-PT(ProTx-II 和 TTX)和 Nav1.7-HS(HWTX-IV 和 STX)。虽然获得了这些结构,并且基于此推断了野生型的结构,但出于 Nav1.7 的潜在药物作用,需要一个野生型高分辨率的结构为药物筛选和机制研究提供参考和帮助。
2022 年 4 月 26 日,颜宁教授团队与西湖大学申怀宗研究员课题组合作解析了 Nav1.7 高分辨率结构,以 High-resolution structures of human Nav1.7 reveal gating modulation through a-p helical transition of S6IV 为题发表在 Cell Reports 杂志。该研究报道了野生型 Nav1.7 和 Nav1.7(E406K)的冷冻电镜结构,分辨率分别为 2.2Å 和 3.5Å。与最新的与 HWTX-IV 或 ProTX-II 结合的 Nav1.7(E406K)结构相比,揭示了这些毒素通过 S6IV 段的 a 到 π 转变对通道门控进行变构调节。 在这一文章中,研究人员报告了野生型(WT)Nav1.7 2.2Å 的高分辨率的冷冻电镜结构。Nav1.7-β1-β2 复合物的表达量约为突变体的一半,研究人员使用 40L 的 293F 细胞获得了可以用于冷冻的蛋白样品,最终,从 78.5 万多个粒子中获得了总分辨率为 2.2Å 的电镜结构。
在这一文章中,研究人员报告了野生型(WT)Nav1.7 2.2Å 的高分辨率的冷冻电镜结构。Nav1.7-β1-β2 复合物的表达量约为突变体的一半,研究人员使用 40L 的 293F 细胞获得了可以用于冷冻的蛋白样品,最终,从 78.5 万多个粒子中获得了总分辨率为 2.2Å 的电镜结构。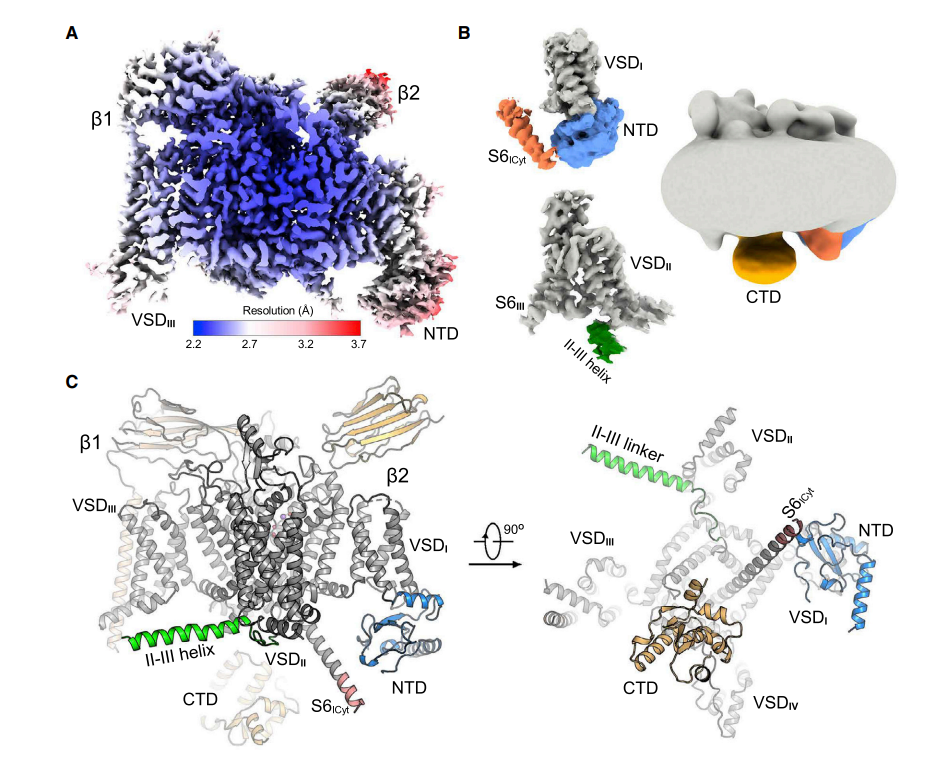
在目前的分辨率下,研究者建立了一个可靠的 NTD 结构域的原子模型。此外,解决了几个细胞内片段,包括重复序列 I 中 S6 片段(S6I)之后的额外螺旋片段、通过柔性连接体连接到 S6II 的反螺旋,以及可能属于羧基末端结构域(CTD)的密度团。
通过对比发现,研究人员对比发现 Nav1.7 和 Nav1.4 的构象非常接近,研究人员还观察到 WT Nav1.7 的胞内门区存在一个 GDN,这与已报道的 Nav1.7-PT 和 Nav1.7-HS 的结果相矛盾。紧接着研究人员比较了以前和现在的结果,以寻找这种差异的分子基础。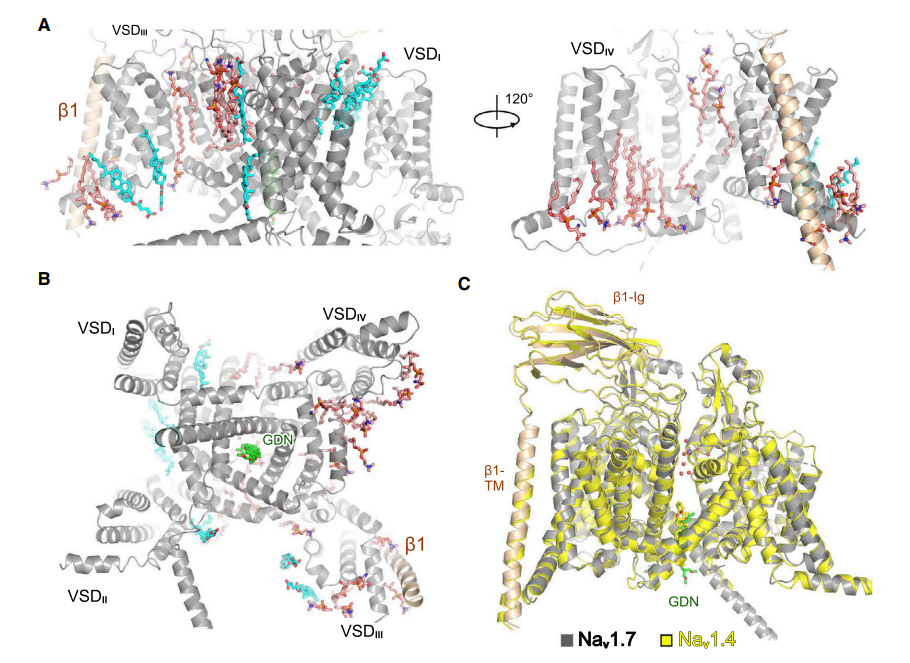
已报道的 Nav1.7-HS/PT 的 EM 图在跨膜域的大部分区域显示良好的质量,但 S6IV 的最后四个螺旋转弯的局部密度更差,根据推断,这种局部不良的分辨率的可能是由于构象异质性造成的,这在最初的研究中没有成功地分类。
Nav1.7-HS 的两个构象在 Val1751 开始偏离。其中一个构象,在 3.1-Å 分辨率,共享与 Nav1.7-WT 相同的结构,其中整个 S6IV 是由螺旋转弯组成的,它们之后被称为 HS-a 和 WT-a。在 Nav1.7-HS 的 3.0-Å 结构中,由 1751VVVN1754 组成的螺旋转弯为 π 型。Nav1.7-PT 也是如此,研究人员将其命名为 HS-π 和 PT-π。
总的来说,通过对 Nav1.7 在毒素缺失和存在情况下的结构进行系统性的比较,发现在与 GMT(gating modifier toxins)结合时 PD 构像发生了远程变构调节。在 π 型结构中,细胞内孔道进一步收缩,一个窗口关闭,IFM 结合位点被重构。与 HWTX-IV 结合的所有 4 个 S6 片段的伴随结构转移,为 PD 中不同重复序列之间的构象耦合提供了一条清晰的路径。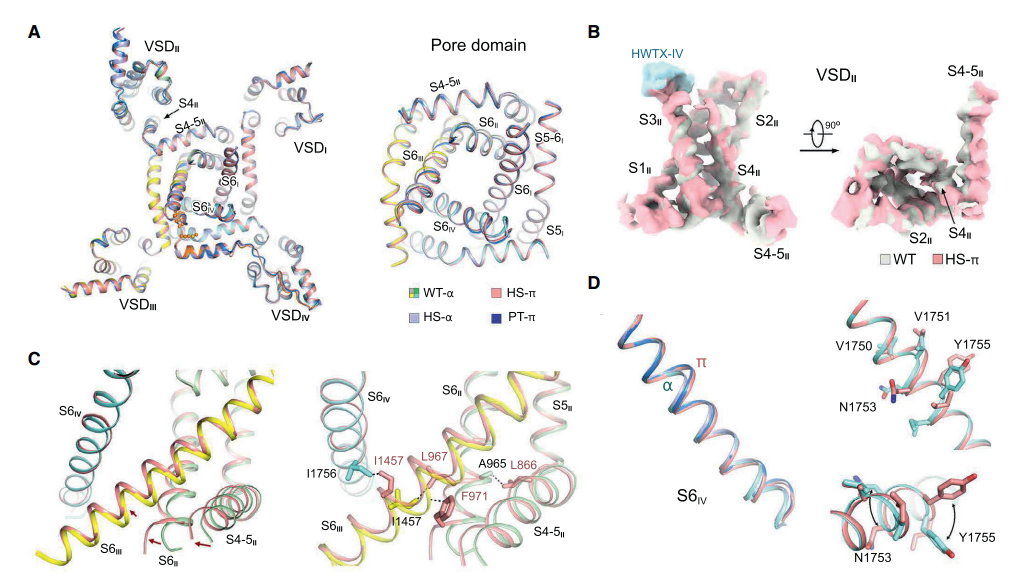
通过 S6IV 简单的 a-π 跃迁,细胞内门脉的耦合、IFM 结合位点的形成以及对开窗的调节都是有趣的,但并不罕见。最近的结构进展表明,a-π 跃迁在电压门控离子通道(VGICs)中很常见。在比较昆虫 NavPaS 和兔子 Cav1.1 的结构时,首先注意到 S6 节段的 a 和 π 形态的差异,它们在 PD 构像上表现出明显的差异。除了单链真核生物 Nav 和 Cav 通道外,在多个 TRP 通道结构中也观察到 a-π 转变参与通道闸门。尽管有这些结构特征,这一次级结构转换的功能相关性仍有待进一步研究。
值得注意的是,在高压激活(HVA)的 Cav1 和 Cav2 通道中也观察到了类似的细胞质结构。尽管缺乏 IFM 基序,HVA Cav 通道包含一个短的 III-IV 螺旋,在 S6III 之后,并与 CTD 发生交互作用。事实上,在昆虫通道 NavPaS 中,III-IV 螺旋与 CTD 的共折叠是保守的,其构象与其他所有导航通道结构不同。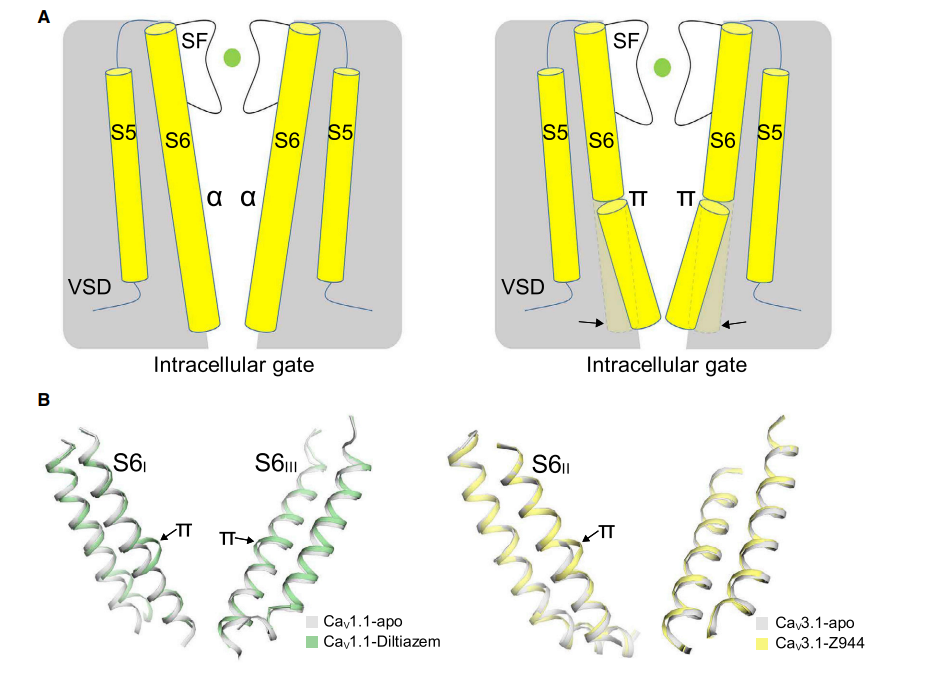
在 Nav 和 Cav 通道中,细胞质螺旋对于重复序列 III 和 IV 是保守的,而重复序列 I 和 II 中的螺旋则被交换。在 Cav2.2 中,S6I 特别长,直接延伸到细胞质中,而 S6I 后面是一个名为 AID 的横向螺旋,是 Nav1.7 中 II-III 螺旋的补充。在 Cav2.2 中,AID 螺旋和 S6II 延伸的胞质片段与辅助 β 亚基相互作用。Nav 通道重复序列 I 和 II 中的这些胞质螺旋是否与未表征的调控蛋白相关还有待研究。
总之,高分辨率的人类 Nav1.7 的结构为进一步的生物物理和计算分析提供了一个准确的模板,这将有助于未来药物的开发。