RNAi的作用机制及其放大效应
互联网
- 相关专题
- RNA干扰技术
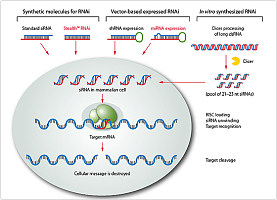
RNAi 的作用机制
RNAi 的作用体现在两种水平上
①转录 水平。RNA 可以介导DNA 的甲基化(RNA-directed DNA methylation ,RdDM) 最早发现于一种植物 的类病毒系统。dsRNA(double-strand RNA) 被降解成21~23 个核苷酸 长的小片段RNA 时,这些小的RNA 分子 在细胞核 可以诱发同源序列的DNA 甲基化 。这种序列特异性的甲基化的信号与RNA-DNA 结合有关 。当dsRNA 含有与启动子同源的序列,即可使同源靶启动子序列甲基化,从而使靶启动子失去功能,导致下游基因沉默。非致病病毒的造成转录 水平基因沉默相关的RdDM最先发现于以菜花样花叶病毒,其35S 启动子转录得到含有胭脂碱合成酶启动子(NOSpro) 序列的NOSproRNA。在转基因烟草中,据报告未腺苷酸化的NOSpro dsRNA可以诱导同源NOSpro DNA 的甲基化,并使拷贝在转录水平上反式失活。在链胞霉属,DNA 的甲基化可能是异染色质的siRNA 介导的组蛋白H3 K9 的甲基化,是由H3 甲基转移酶催化的 。
②转录后水平。⑴特异切割dsRNA 的RNA 酶Ⅲ家族核酸酶Dicer 依赖ATP 切割dsRNA ,将其分解成具有2 个核苷酸的3′悬端(overhang) 的19~21b 小片段的双链siRNA。⑵RISC(RNA 诱导的沉默复合物RNA-induced silence complex) 识别并降解mRNA。RISC 是一种蛋白-RNA 效应器核酸酶复合物。双链siRNA 为RISC 的重要组分,它依赖ATP 解旋导致RISC活化,然后通过Waston-Crick 碱基配对识别底物mRNA 并与之结合,并自siRNA 的3′端将mRNA 切割成小于12nt 的片段使其降解 。对一些人组织因子的试验表明,几个不同的siRNA 可以攻击同一个mRNA 的不同位点。只有一部分siRNA 会导致显著的基因沉默,这说明在人mRNA 上的siRNA 结合位点可能很少,且不活跃的siRNA 与活跃的siRNA 可逆性竞争。
miRNA
Dicer 酶产生两类功能特异的小分子 RNA:siR2NA 和miRNA(micro-RNA)。Tuschl 等在从果蝇 胚胎提取物的长的dsRNA 处理得到siRNA 的同时,得到了16 种20—23个核苷酸大小的短的单链RNA。这些RNA 由果蝇 基因组 编码并在0—2 周的胚胎内表达。它们具有潜在的调节基因表达 的作用,被称为miRNA 。miRNA 约21 个核苷酸大小,为单链结构,大多数miRNA 与底物mRNA 不完全配对结合,它们并不改变miRNA 的稳定性,而是通过抑制翻译来使基因沉默,但目前至少发现一种植物miRNA(miR171) 与底物mRNA完全配对,这提示了miRNA 通过RNAi 途径调节基因表达 的可能性 。与siRNA 不同的是,由于miRNA 为单链结构,其作用不需要ATP 的存在。
RNAi 的放大效应
由于RNAi 的机制在生物体的效应极其显著,有研究提示,RNAi 通路 应该有相应的放大步骤。放大效应可能的途径:①通过复制dsRNA 或者siRNA 进行。Nishikura 提出“过渡的RNAi (transitive RNAi) ”的机制,即由原先的靶序列的上游序列延伸形成新的siRNA,继续降解同源的基因家族成员或者切割mRNA 。②多轮的RISC 效应。③RdRP(RNA 诱导的RNA 多聚酶) 的作用:植物和新秀丽小杆线虫的dsRNA 诱导的沉默都需要与RdRP 相似的蛋白质的参与 。RNAi 一般在体细胞发挥作用,在ego21 表达的生殖细胞一种称为“随机降解PCR ”的放大模型却提示RdRP 通过siRNA 引导链识别并结合mRNA,产生一种双链RNA 的Dicer酶底物,以产生更多的siRNA。