RNAi技术的研究进展与发展前景
互联网
(RNA interference,RNAi)是1998年由Fire等在线虫中发现的一种转录后的基因沉默 (Posttranscriptional gene silencing,PTGS)机制。双链RNA(double-stranded RNA,dsRNA)能特异地抑制或沉默目的基因表达,产生如同目的基因突变的缺陷表型,这种由dsRNA介导的基因阻抑作用被称为RNAi。1990年,美国和荷兰的两个转基因植物实验组,在矮牵牛中发现的一种转基因能同时抑制相应的内源基因以及自身表达的基因沉默现象,当时将这种现象称为共抑制。到1994 年,由Cogni等在真菌中发现转录后的基因沉默现象,在野生型粗糙链胞霉中转入胡萝卜素基因 albino 1或albino 3时,发现在部分实验中,内源性al 1或al 3基因的表达水平反而减弱,称此现象为基因压制(quelling)。1998年,Fire等在线虫中发现了RNAi现象,并揭示了SuGuo发现正义 RNA对基因表达也有抑制作用的原因,认为SuGuo发现的这种现象是由于体外转录制备的RNA中污染了微量的双链RNA,而且dsRNA能比反义RNA或正义RNA更有效地抑制基因的表达,把由 RNA引起的基因表达的抑制称为RNA干涉。
最初普遍认为共抑制、基因压制以及RNAi是机制完全不同的基因抑制现象。但经过科研人员的不断研究,发现在共抑制、真菌中的基因压制以及RNAi现象之间存在着密切的联系。都是由 RNA引起的转录后的基因沉默,可能有共同的
生物
学意义和相似的作用机制。但是共抑制与 RNAi并不是完全相同,在植物的共抑制中,dsRNA不仅能引起转录后的基因沉默,而且还能引起转录水平的沉默,其可能机制是dsRNA能引起染色质的重组或甲基化而改变其内源基因的序列。因此,在植物共抑制中还存在RNAi以外的由RNA指导的DNA甲基化,而引起转录水平抑制的机制。dsRNA能特异地抑制目的基因的表达,其广泛存在各种有机体中,包括线虫、果蝇、涡虫、水螅、锥虫、真菌、植物以及哺乳动物。
1 RNAi的特征
RNAi是发生在转录后水平的基因沉默。
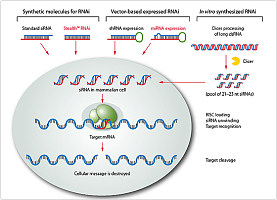
dsRNA具有很高的特异性,能特异地将与其同源的mRNA降解。Andrew在植物中用3种转基因诱导的转录后的基因沉默(PTGS)都检测到有约25 nt(nucleotide)长的siRNA(short interference RNA)。Shi Chenyang等从已转染dsRNA的未分化的胚胎干细胞、卵母细胞以及老鼠胚胎细胞的细胞质提取物中都发现有21~23 nt长的siRNA。这些siRNA可能具有指导核酸酶特异地识别靶mRNA 而将其降解的功能,21~23 nt以下的片段还没有发现,由于这些片段不稳定,易被胞内其他核酸酶降解。
极低浓度的dsRNA就能完全抑制基因的表达。这可能由于RNA存在复制或由于dsRNA具有很强的催化功能。
dsRNA抑制效应具有传递性。Timmons等用含目的基因表达dsRNA载体大肠杆菌喂养线虫, RNA从肠中吸收,但在体细胞及生殖细胞中都有分布表达。此外,RNAi不仅发生在亲代动物本身,其子代伴随基因表达过程也产生了强烈而特异的抑制效应。细胞的这种抑制能力还能在细胞与细胞之间通过胞间连丝传递,甚至可以通过植物的维管组织在整个植物体中传递。Jorgenese等将有共抑制现象的植物作为砧木,没有抑制现象的植物作为接穗,一段时间以后在接穗中产生了共抑制现象。说明有某种信号分子通过植物的维管系统进行传递,由于这种系统获得的沉默具有序列特异性的特点,这种信号分子可能是一种RNA与蛋白质的复合体。