光合作用基础知识讲座系列二:叶绿体和光合色素
互联网
一、叶绿体
叶片是光合作用的主要器官,而叶绿体(chloroplast,chlor)是光合作用最重要的细胞器。
(一)叶绿体的发育、形态及分布
1.发育 高等植物的叶绿体由前质体(proplastid)发育而来,前质体是近乎无色的质体,它存在于茎端分生组织中。当茎端分生组织形成叶原基时,前质体的双层膜中的内膜在若干处内折并伸入基质扩展增大,在光照下逐渐排列成片,并脱离内膜形成囊状结构的类囊体,同时合成叶绿素,使前质体发育成叶绿体。幼叶绿体能进行分裂。
2.形态 高等植物的叶绿体大多呈扁平椭圆形,每个细胞中叶绿体的大小与数目依植物种类、组织类型以及发育阶段而异。一个叶肉细胞中约有10至数百个叶绿体,其长3~7μm,厚2~3μm。
3.分布 叶肉细胞中的叶绿体较多分布在与空气接触的质膜旁,在与非绿色细胞(如表皮细胞和维管束细胞)相邻处,通常见不到叶绿体。这样的分布有利于叶绿体同外界进行气体交换。
4.运动 叶绿体在细胞中不仅可随原生质环流运动,而且可随光照的方向和强度而运动。在弱光下,叶绿体以扁平的一面向光以接受较多的光能;而在强光下,叶绿体的扁平面与光照方向平行,不致吸收过多强光而引起结构的破坏和功能的丧失。
(二)叶绿体的基本结构
叶绿体是由叶绿体被膜、基质和类囊体三部分组成。
1.叶绿体被膜(chloroplast envelope) 叶绿体被膜由两层单位膜组成,两膜间距5~10nm。被膜上无叶绿素,它的主要功能是控制物质的进出,维持光合作用的微环境。外膜(outer membrane)为非选择性膜,分子量小于10000的物质如蔗糖、核酸、无机盐等能自由通过。内膜(inner membrane)为选择透性膜,CO2、O2、H2O可自由通过;Pi、磷酸丙糖、双羧酸、甘氨酸等需经膜上的运转器(translocator)才能通过;蔗糖、C5`C7糖的二磷酸酯、NADP+、PPi等物质则不能通过。
2.基质及内含物 被膜以内的基础物质称为基质(stroma),基质以水为主体,内含多种离子、低分子的有机物,以及多种可溶性蛋白质等。基质是进行碳同化的场所,它含有还原CO2与合成淀粉的全部酶系,其中1,5-二磷酸核酮糖羧化酶/加氧酶(ribulose1,5bisphosphate carboxylase/oxygenase,Rubisco)占基质总蛋白的一半以上。此外,基质中含有氨基酸、蛋白质、DNA、RNA、脂类(糖脂、磷脂、硫脂)、四吡咯(叶绿素类、细胞色素类)和萜类(类胡萝卜素、叶醇)等物质及其合成和降解的酶类,还含有还原亚硝酸盐和硫酸盐的酶类以及参与这些反应的底物与产物,因而在基质中能进行多种多样复杂的生化反应。
基质中有淀粉粒(starch grain)与质体小球(plastoglobulus),它们分别是淀粉和脂类的贮藏库。将照光的叶片研磨成匀浆离心,沉淀在离心管底部的白色颗粒就是叶绿体中的淀粉粒。质体小球又称脂质球或亲锇颗粒,在叶片衰老时叶绿体中的膜系统会解体,此时叶绿体中的质体小球也随之增多增大。
3.类囊体 类囊体(thylakoid)是由单层膜围起的扁平小囊,膜厚度5~7nm,囊腔(lumen)空间为10nm左右,片层伸展的方向为叶绿体的长轴方向。类囊体分为二类:一类是基质类囊体(stroma thylakoid),又称基质片层(stroma lamella),伸展在基质中彼此不重叠;另一类是基粒类囊体(grana thlylakoid),或称基粒片层(grana lamella),可自身或与基质类囊体重(granum)。片层与片层互相接触的部分称为堆叠区(appessed region),其他部位则为非堆叠区(nonappressed region)。
(三)类囊体膜上的蛋白复合体
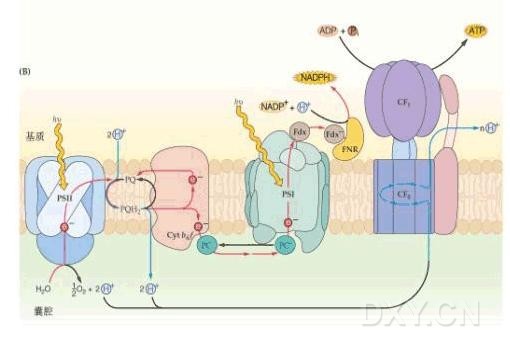
类囊体膜上含有由多种亚基、多种成分组成的蛋白复合体,主要有四类,即光系统Ⅰ(PSI)、光系统Ⅱ(PSⅡ)、Cytb6/f复合体和ATP酶复合体(ATPase),它们参与了光能吸收、传递与转化、电子传递、H+输送以及ATP合成等反应。由于光合作用的光反应是在类囊体膜上进行的,所以称类囊体膜为光合膜(photosynthetic membrane)。这四类蛋白复合体在类囊体膜上的分布大致是:PSⅡ主要存在于基粒片层的堆叠区,PSⅠ与ATPase存在于基质片层与基粒片层的非堆叠区,Cytb6/f复合体分布较均匀。PSⅡ中放氧复合体(oxygen-evolving complex,OEC)在膜的内表面,PSⅡ的原初供体位于膜内侧,原初受体靠近膜外侧。质体醌(plastoquinone,PQ)可以在膜的疏水区内移动。Cytb6/f复合体在膜的疏水区。PSⅠ的电子供体PC在膜的内腔侧,而PSⅠ还原端的Fd、FNR在膜的外侧。蛋白复合体及其亚基的这种分布,有利于电子传递、H+的转移和ATP合成。
二、光合色素
在光合作用的反应中吸收光能的色素称为光合色素,主要有三种类型:叶绿素、类胡萝卜素和藻胆素。高等植物中含有前两类,藻胆素仅存在于藻类中。
(一)光合色素的结构和性质
1.叶绿素 叶绿素(chlorophyll)是使植物呈现绿色的色素,约占绿叶干重的1%。植物的叶绿素包括a、b、c、d四种。高等植物中含有a、b两种,叶绿素c、d存在于藻类中,而光合细菌中则含有细菌叶绿素(bacteriochlorophyll)。叶绿素a(Chl a)呈蓝绿色,叶绿素b(Chl b)呈黄绿色,分子量分别为892和906,叶绿素是双羧酸的酯,其中一个羧基被甲醇所酯化,另一个被叶绿醇所酯化,它们的分子式可以写成:
叶绿素a与b的分子式很相似,不同之处是叶绿素a比b多两个氢少一个氧。两者结构上的差别仅在于叶绿素a的B吡咯环上一个甲基(-CH3)被醛基(-CHO)所取代。
叶绿素分子含有一个卟啉环(porphyrin ring)的“头部”和一个叶绿醇(植醇,phytol)的“尾巴”。卟啉环由四个吡咯环与四个甲烯基(-CH=)连接而成,它是各种叶绿素的共同基本结构。卟啉环的中央络合着一个镁原子,镁偏向带正电荷,而与其相联的氮原子则带负电荷,因而“头部”有极性,是亲水的。另外还有一个含羰基的同素环(含相同元素的环),其上一个羧基以酯键与甲醇相结合。环D上有一个丙酸侧链以酯键与叶绿醇相结合,叶绿醇是由四个异戊二烯单位所组成的双萜,是亲脂的,能伸入类囊体的拟脂层,故叶绿素能定向排列。
卟环上的共轭双键和中央镁原子容易被光激发而引起电子的得失,这决定了叶绿素具有特殊的光化学性质。 卟啉环中的镁可被H+、Cu2+、Zn2+ 等所置换。当为H+所置换后,即形成褐色的去镁叶绿素(pheophytin,Pheo)。去镁叶绿素中的H+ 再被Cu2+取代,就形成铜代叶绿素,颜色比原来的叶绿素更鲜艳稳定。根据这一原理可用醋酸铜处理来保存绿色标本。 叶绿素是一种酯,因此不溶于水。
通常用含有少量水的有机溶剂如80%的丙酮,或者95%乙醇,或丙酮∶乙醇∶水=4.5∶4.5∶1的混合液来提取叶片中的叶绿素,用于测定叶绿素含量。之所以要用含有水的有机溶剂提取叶绿素,这是因为叶绿素与蛋白质结合很牢,需要经过水解作用才能被提取出来。
2.类胡萝卜素 类胡萝卜素(carotenoid)是由8个异戊二烯形成的四萜,含有一系列的共轭双键,分子的两端各有一个不饱和的取代的环己烯,也即紫罗兰酮环,它们不溶于水而溶于有机溶剂。类胡萝卜素包括胡萝卜素(carotene,C40H56O2)和叶黄素(xanthophyll, C40H56O2)。前者呈橙黄色,后者呈黄色。胡萝卜素是不饱和的碳氢化合物,有α、β、γ三种同分异构体,其中以β胡萝卜素在植物体内含量最多。
叶黄素是由胡萝卜素衍生的醇类,也叫胡萝卜醇(carotenol),通常叶片中叶黄素与胡萝卜素的含量之比约为2:1。
类胡萝卜素除了有吸收传递光能的作用外,还可在强光下逸散能量,如β-胡萝卜素就是单线态分子氧(1O2)的猝灭剂,具有使叶绿素免遭伤害的光保护作用(photoprotection)。
一般来说,叶片中叶绿素与类胡萝卜素的比值约为3∶1,所以正常的叶子总呈现绿色。秋天或在不良的环境中,叶片中的叶绿素较易降解,数量减少,而类胡萝卜素比较稳定,所以叶片呈现黄色。类胡萝卜素总是和叶绿素一起存在于高等植物的叶绿体中,此外也存在于果实、花冠、花粉、柱头等器官的有色体中。
3.藻胆素(phycobilin) 仅存在于红藻和蓝藻中,主要有藻红蛋白(phycoerythrin)、藻蓝蛋白(phycocyanin)和别藻蓝蛋白(allophycocyanin)三类,前者呈红色,后两者呈蓝色。它们的生色团与蛋白以共价键牢固地结合。藻胆素分子中的四个吡咯环形成直链共轭体系,不含镁也没有叶绿醇链。藻胆素也有收集光能的功能。
由于类胡萝卜素和藻胆素吸收的光能能够传递给叶绿素用于光合作用,因此它们被称为光合作用的辅助色素(accessory photosynthetic pigments)。
(二)光合色素的吸收光谱
用分光光度计能精确测定光合色素的吸收光谱(absorption spectrum)(图4-6)。叶绿素最强的吸收区有两处:波长640~660nm的红光部分和430~450nm的蓝紫光部分。叶绿素对橙光、黄光吸收较少,尤以对绿光的吸收最少,所以叶绿素的溶液呈绿色。
叶绿素a和叶绿素b的吸收光谱很相似,但也稍有不同:叶绿素a在红光区的吸收峰比叶绿素b的高,而蓝光区的吸收峰则比叶绿素b的低,也就是说,叶绿素b吸收短波长蓝紫光的能力比叶绿素a强。
一般阳生植物叶片的叶绿素a/b比值约为3∶1,而阴生植物的叶绿素a/b比值约为2.3∶1。叶绿素b含量的相对提高就有可能更有效地利用漫射光中较多的蓝紫光,所以叶绿素b有阴生叶绿素之称。
类胡萝卜素的吸收带在400~500nm的蓝紫光区,它们基本不吸收红.橙.黄光,从而呈现橙黄色或黄色.
藻蓝蛋白的吸收光谱最大值是在橙红光部分,而藻红蛋白则是在绿光部分。
植物体内不同光合色素对光波的选择吸收是植物在长期进化中形成的对生态环境的适应,这使植物可利用各种不同波长的光进行光合作用。
(三)叶绿素的生物合成及其与环境条件的关系
1.叶绿素的生物合成 叶绿素的合成是在前质体或叶绿体中在一系列酶的作用下形成的。合成叶绿素分子中的吡咯环的起始物质是δ-氨基酮戊酸(δ-aminolevulinic acid,ALA),在高等植物中ALA由谷氨酸或α-酮戊二酸转化而来。叶绿素合成过程见图4-7。图中(1)2分子ALA脱水缩合形成1分子具有吡咯环的胆色素原;(2)、(3)4分子胆色素原脱氨基缩合形成1分子尿卟啉原Ⅲ,合成过程按A→B→C→D环的顺序进行;(4)尿卟啉原Ⅲ四个乙酸链脱羧形成具有四个甲基的粪卟啉原Ⅲ(5)、(6)粪卟啉原Ⅲ再脱羧、脱氢,氧化形成原卟啉Ⅸ;(7)原卟啉Ⅸ与镁结合形成Mg原卟啉Ⅸ;(8)Mg原卟啉Ⅸ的一个羧基被甲基所酯化;(9)、(10)在Mg-原卟啉Ⅸ上形成第五个环,接着B环上的-CH2=CH2侧链被还原为-CH2-CH3,即形成原叶绿酸酯;(11)经光还原,D环上接受NADPH提供的H,形成叶绿酸酯a,这一过程是在一种色素蛋白复合体上进行的;(12)D环上的丙酸为20个C的牻牛儿基牛牻牛儿基焦磷酸酯化,后者由NADPH提供H还原成叶绿醇,这样就形成了叶绿素a。叶绿素b则是由叶绿素a氧化形成的。

图4-7 叶绿素a的生物合成过程
起始物是谷氨酸,之后为5-氨基酮戊酸,两分子的ALA缩合形成胆色素原(PBG),4分子PBG相互连结形成原中卟啉IX.原卟啉IX与Mg结合形成Mg-原卟啉原IX,光下E环的环化形成,D环的还原作用和叶绿醇尾部的连接完成了整个合成过程,合成过程中的许多步骤在图中已省略
2.影响叶绿素形成的条件
(1)光 光是影响叶绿素形成的主要条件。从原叶绿素酸酯转变为叶绿酸酯需要光,而光过强,叶绿素又会受光氧化而破坏。黑暗中生长的幼苗呈黄白色,遮光或埋在土中的茎叶也呈黄白色。这种因缺乏某些条件而影响叶绿素形成,使叶子发黄的现象,称为黄化现象(etiolation)。
也有例外情况,例如藻类、苔藓、蕨类和松柏科植物在黑暗中可以合成叶绿素,其数量当然不如在光下形成的多;柑橘种子的子叶及莲子的胚芽在无光照的条件下也能形成叶绿素,推测这些植物中存在可代替可见光促进叶绿素合成的生物物质。
(2)温度 叶绿素的生物合成是一系列酶促反应,受温度影响。叶绿素形成的最低温度约2℃,最适温度约30℃,最高温度约40℃。秋天叶子变黄和早春寒潮过后秧苗变白,都与低温抑制叶绿素形成有关。高温下叶绿素分解大于合成,因而夏天绿叶蔬菜存放不到一天就变黄;相反,温度较低时,叶绿素解体慢,这也是低温保鲜的原因之一。
(3)营养元素 叶绿素的形成必须有一定的营养元素。氮和镁是叶绿素的组成成分,铁、锰、铜、锌等则在叶绿素的生物合成过程中有催化功能或其它间接作用。因此,缺少这些元素时都会引起缺绿症(chlorosis),其中尤以氮的影响最大,因而叶色的深浅可作为衡量植株体内氮素水平高低的标志。
(4)氧 缺氧能引起Mg-原卟啉IX或Mg-原卟啉甲酯的积累,影响叶绿素的合成。
(5)水 缺水不但影响叶绿素生物合成,而且还促使原有叶绿素加速分解,所以干旱时叶片呈黄褐色。