酵母双杂交技术及其在蛋白质组研究中的应用
互联网
作为后基因组时代出现的新兴研究领域之一,蛋白质组学(proteomics)正受到越来越多的关注。蛋白质组学的研究目标是对机体或细胞的所有蛋白质进行鉴定和结构功能分析。蛋白质组学的研究不局限任何特定的方法。
高分辨率的蛋白质分离技术如二维凝胶电泳和高效液相层析,经典的蛋白质鉴定方法如氨基酸序列分析等,现代质谱技术,基因组学研究的各种手段,现代计算机信息学和计算机网络通讯技术等等,任何可用于蛋白质研究的技术手段,蛋白质组学都可能会采用。它体现的是一个开放的思维和研究方式。
蛋白质-蛋白质的相互作用是细胞生命活动的基础和特征。这种千变万化的相互作用以及由此形成的纷繁复杂的蛋白质联系网络同样也是蛋白质组学的研究内容,相应的工作也已经开展。
酵母双杂交系统(yeast two-hybrid system)自建立以来已经成为分析蛋白质相互作用的强有力的方法之一。
该方法在不断完善,如今它不但可用来在体内检验蛋白质间的相互作用,而且还能用来发现新的作用蛋白质,在对蛋白质组中特定的代谢途径中蛋白质相互作用关系网络的认识上发挥了重要的作用。实验表明双杂交技术在蛋白质组学上的应用是成功的。本文将就双杂交技术的产生、发展及其在蛋白质组研究方面的初步应用作一介绍。
1 蛋白质组学的产生背景
基因组研究自从开展以来已经取得了举世瞩目的成就。在过去几年中,已经陆续完成了包括大肠杆菌、酿酒酵母等十多种结构比较简单的生物的基因组DNA的全序列分析。线虫(C.elegans)的基因组DNA测序工作已基本完成。
规模更为庞大的人类基因组计划预期在下一世纪的前几年(2003~2005年)也将完成全部基因组DNA的序列分析。这些进展是非常令人振奋的。但是也随之产生了新问题。大量涌出的新基因数据迫使我们不得不考虑这些基因编码的蛋白质有什么功能这个问题。
不仅如此,在细胞合成蛋白质之后,这些蛋白质往往还要经历翻译后的加工修饰。
也就是说,一个基因对应的不是一种蛋白质而可能是几种甚至是数十种。包容了数千甚至数万种蛋白质的细胞是如何运转的?或者说这些蛋白质在细胞内是怎样工作、如何相互作用、相互协调的?这些问题远不是基因组研究所能回答得了的。正是在此背景下,蛋白质组学(proteomics)应运而生。
蛋白质组(proteome)一词是马克.威尔金斯(Marc Wilkins)最先提出来的,最早见诸于1995年7月的“Electrophoresis”杂志上,它是指一个有机体的全部蛋白质组成及其活动方式。蛋白质组研究虽然尚处于初始阶段,但已经取得了一些重要进展。
当前蛋白质组学的主要内容是,在建立和发展蛋白质组研究的技术方法的同时,进行蛋白质组分析。对蛋白质组的分析工作大致有两个方面。一方面,通过二维凝胶电泳得到正常生理条件下的机体、组织或细胞的全部蛋白质的图谱,相关数据将作为待检测机体、组织或细胞的二维参考图谱和数据库。
一系列这样的二维参考图谱和数据库已经建立并且可通过联网检索。二维参考图谱建立的意义在于为进一步的分析工作提供基础。
蛋白质组分析的另一方面,是比较分析在变化了的生理条件下蛋白质组所发生的变化。如蛋白质表达量的变化,翻译后修饰的变化,或者可能的条件下分析蛋白质在亚细胞水平上的定位的改变等。关于蛋白质组学的介绍可参阅文献。
细胞或组织的蛋白质不是杂乱无章的混合物,蛋白质间的相互作用、相互协调是细胞进行一切代谢活动的基础。
蛋白质间的相互作用及作用方式同样也是蛋白质组研究所面临的问题。研究蛋白质间的相互作用有多种方法,常用的如酵母双杂交系统、亲和层析、免疫沉淀、蛋白质交联等。其中,酵母双杂交系统是当前发展迅速、应用广泛的主要方法。
2 酵母双杂交系统的建立与发展
双杂交系统的建立得力于对真核生物调控转录起始过程的认识。细胞起始基因转录需要有反式转录激活因子的参与。
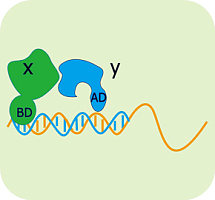
80年代的工作表明,转录激活因子在结构上是组件式的(modular),即这些因子往往由两个或两个以上相互独立的结构域构成,其中有DNA结合结构域(DNA binding domain,简称为DB)和转录激活结构域(activation domain,简称为AD),它们是转录激活因子发挥功能所必需的。单独的DB虽然能和启动子结合,但是不能激活转录。
而不同转录激活因子的DB和AD形成的杂合蛋白仍然具有正常的激活转录的功能。如酵母细胞的Gal4蛋白的DB与大肠杆菌的一个酸性激活结构域B42融合得到的杂合蛋白仍然可结合到Gal4结合位点并激活转录。
Fields等人的工作标志双杂交系统的正式建立。他们以与调控SUC2基因有关的两个蛋白质Snf1和Snf2为模型,将前者与Gal4的DB结构域融合,另外一个与Gal4的AD结构域的酸性区域融合。由DB和AD形成的融合蛋白现在一般分别称之为“诱饵”(bait)和“猎物”或靶蛋白(prey or target protein)。
如果在Snf1和Snf2之间存在相互作用,那么分别位于这两个融合蛋白上的DB和AD就能重新形成有活性的转录激活因子,从而激活相应基因的转录与表达。这个被激活的、能显示“诱饵”和“猎物”相互作用的基因称之为报道基因(reporter gene)。
通过对报道基因表达产物的检测,反过来可判别作为“诱饵”和“猎物”的两个蛋白质之间是否存在相互作用。在此Fields等人采用编码β-半乳糖苷酶的LacZ作为报道基因,并且在该基因的上游调控区引入受Gal4蛋白调控的GAL1序列。
这个改造过的LacZ基因被整合到酵母染色体URA3位上。而酵母的GAL4基因和GAL80基因(Gal80是Gal4的负调控因子)被缺失,从而排除了细胞内源调控因子的影响。已经知道在Snf1和Snf2之间存在相互作用。
结果发现只有同时转化了Snf1和Snf2融合表达载体的酵母细胞才有β-半乳糖苷酶活性,单独转化其中任何一个载体都不能检测出β-半乳糖苷酶活性。
目前发展起来的各种双杂交系统大多是以Fields等人建立的系统为基础的。这些新系统主要对报道基因、“诱饵”表达载体以及“猎物”表达载体等做了一些改进。其中一个重要改进是引入额外的报道基因,如广泛采用的HIS3基因。
经过改造带有HIS3报道基因的酵母细胞,只有当HIS3被启动表达才能在缺乏组氨酸的选择性培养基上生长。HIS3报道基因的转录表达是由“诱饵”和“猎物”的相互作用所启动的。大多数双杂交系统往往同时使用两个甚至三个报道基因,其中之一是LacZ。
这些改造后的基因在启动子区有相同的转录激活因子结合位点,因此可以被相同的转录激活因子(如上述的Gal4蛋白)激活。通过这种双重或多重选择既提高了检测灵敏度又减少了假阳性现象。其他还有针对“诱饵”或“猎物”表达载体等所作的改进,这里不一一详述。
在双杂交鉴定过程中要经过两次转化,这个工作量是相当大的,特别是寻找新的作用蛋白质的时候尤其如此。而且,酵母细胞的转化效率比细菌要低约4个数量级。因此转化步骤就成为双杂交技术的瓶颈。
Bendixen等人通过酵母接合型的引用,避免了两次转化操作,同时又提高了双杂交的效率。
在酵母的有性生殖过程中涉及到两种配合类型: a接合型和α接合型,这两种单倍体之间接合(mating)能形成二倍体,但a接合型细胞之间或α接合型细胞之间不能接合形成二倍体。
根据酵母有性生殖的这一特点,他们将文库质粒转化α接合型酵母细胞,“诱饵”表达载体转化a接合型细胞。然后分别铺筛选平板使细胞长成菌苔(lawn),再将两种菌苔复印到同一个三重筛选平板上,原则上只有诱饵和靶蛋白发生了相互作用的二倍体细胞才能在此平板上生长。
单倍体细胞或虽然是二倍体细胞但DB融合蛋白和AD融合蛋白不相互作用的都被淘汰。
长出来的克隆进一步通过β-半乳糖苷酶活力进行鉴定。这项改进不仅简化了实验操作,而且也提高了双杂交的筛选效率。
在研究蛋白质的结构功能特点、作用方式过程中,有时还要通过突变、加抑制剂等手段破坏蛋白质间的相互作用。针对实际工作中的这种需要,Vidal等人发展了所谓的逆双杂交系统(reverse two-hybrid system)。这项技术的关键是报道基因URA3的引入。URA3基因在这里起到了反选择的作用,它编码的酶是尿嘧啶合成的关键酶。该酶能把5-氟乳清酸(5-FOA)转化成对细胞有毒的物质。Vidal等人通过改造在URA3基因的启动子内引入Gal4的结合位点。这个改造的酵母菌株在缺乏尿嘧啶的选择性培养基上只有当“诱饵”和“猎物”相互作用激活URA3基因的表达才能生长。在含有5-FOA的完全培养基上“诱饵”和“猎物”的相互作用则抑制细胞的生长。然而如果目的蛋白,即与DB或AD融合的蛋白质发生了突变或者由于外加药物的干扰不再相互作用,URA3基因不表达,则细胞能在含有5-FOA的完全培养基上生长。通过这种方法,Vidal等人筛选到了转录因子E2F1的突变物,这些突变物仍然能结合视网膜母细胞瘤蛋白RB,但是丧失了同另外一种称为DP1蛋白的结合能力。结果得到了体外结合实验的验证。通过对这些突变蛋白基因的测序,他们发现了新的E2F1同DP1结合的位点。
以上介绍的酵母双杂交系统都是建立在对RNA聚合酶II的激活的基础上的。这意味着双杂交过程是在细胞核内完成的。然而许多待研究的蛋白质是膜蛋白,它们需定位到膜上之后才能表现正常的生物功能,与其他蛋白质起作用。有些真核细胞蛋白还要经过翻译后的加工修饰如二硫键的形成、糖基化、聚异戊二烯基化等。这些加工修饰在正常的细胞核内不可能发生。有些蛋白质本身具有激活转录的活性,它们可不经过双杂交相互作用即能激活报道基因的表达。因此根据需要有人又发展了新的双杂交系统。例如,Aronheim等人设计了一个Sos蛋白介导的双杂交系统,也被作者称为Sos救活系统(Sos recruitment system)。人的鸟苷酸交换因子-Sos蛋白是一种胞质蛋白,而酵母的鸟苷酸交换因子-cdc25蛋白定位在内膜上,可将相关受体信号传导给Ras蛋白。由cdc25突变产生的一种温度敏感突变型细胞不能在36 ℃条件下生长。但是,如果Sos蛋白可以通过某种方式被定位到酵母细胞内膜上,那么它在这种温度敏感突变型细胞中因替代cdc25蛋白的作用,而使酵母恢复在36 ℃条件下生长的能力。在Aronheim等人设计的系统中,待研究的两个蛋白质分别与豆蔻酰基化信号片段和Sos蛋白融合,前一个融合蛋白定位于膜上,如果两个待研究蛋白质之间存在相互作用,这将使Sos也定位到膜上,从而可激活Ras蛋白使酵母在36 ℃下生长。利用该技术Aronheim等人找到了c-Jun的两个新的作用伙伴。
此外还有其他类型的双杂交甚至三杂交系统,本文不再详述。
3 双杂交系统在蛋白质组研究中的应用
双杂交系统在蛋白质组研究上的应用可以说尚处于尝试阶段,这方面的文献报道还很少,但已显示其在分析鉴定细胞内复杂的蛋白质相互作用方面的潜力。这里以两家实验室的工作为例作一介绍。
Fromont-Racine等人以10种功能已知的与mRNA前体剪接有关的蛋白质作起始“诱饵”,从含有约5×106个克隆的酿酒酵母基因组文库中进行第一轮筛选(这些基因组DNA与AD的基因融合构成“猎物”表达载体)。经过对阳性克隆“猎物”DNA的序列分析,他们把这些DNA分成两大类共5项(图1): 编码蛋白质的DNA和非编码蛋白质的DNA,前一类分为四项: A1是在序列上彼此有部分重叠的克隆群。这一项也是首先分析考虑的对象; A2和A3分别是靠近ORF起始密码子的编码蛋白质N-末端的片段和ORF内部的大编码片段,A4是其他的编码序列。这四项优先考虑的顺序是: A1>A2>A3>A4。根据分析把从中得到的4种靶蛋白做成新的“诱饵”进行第二轮筛选。再把从中得到的一种“猎物”做成“诱饵”进行第三轮筛选。如此经过三轮共十五次筛选,他们从中共得到约700个阳性克隆并且对大多数作了部分测序。这些筛选到的靶蛋白中,有9种是已知的mRNA前体剪接因子,5种是新发现的剪接因子,8种是与RNA其他加工过程有关的因子,还有45种与其他的功能有关。另外有9种是转录激活因子,这主要来自假阳性克隆。他们不仅发现了已知剪接因子与其他因子的相互作用,鉴定了一些新的因子,而且建立了这个剪接途径与其他代谢途径的联系。
据此Fromont-Racine等人声称,如果选择不同的细胞代谢或信号转导途径为出发点,通过类似的双杂交实验最终有可能建立酵母细胞完整的蛋白质联系图谱。推而广之,这项技术同样也能用到人类蛋白质组研究中。
Fromont-Racine等人能在较短的时间完成如此大量的筛选、分析工作,一个非常关键的因素是他们采用了上述Bendixen等人建立的通过酵母的有性接合得到双杂交菌株,并且对此技术作了进一步的改进。Bendixen等人是通过把两种接合型细胞同时复印到一个平板上使之接合;而Fromont-Racine则是把两种细胞直接在滤膜上混合然后铺到选择性平板上,因此避免了烦琐的复印操作,使工作效率进一步有所提高。
与此类似,Hudson等人也正在进行酿酒酵母的蛋白质组的研究分析工作。他们把酵母所有的即约6 000多种蛋白质开放读码框(ORF),分别构建成与DB和AD基因融合的表达载体。两类载体分别转化不同接合型酵母菌株。6 000株分别表达不同的AD-ORF即“猎物”的细胞在16张384孔微滴定板上培养,再取少许菌液点在只表达一种BD融合蛋白的细胞形成的菌苔上使两种细胞接合形成二倍体,然后按照常规的双杂交技术鉴定“诱饵”和AD融合蛋白是否发生相互作用(图2)。整个过程主要是在384孔微滴定板上完成的。和Fromont-Racine等人的方法相比,这个方法的优点是操作过程可最大程度地自动化,微滴定板的使用也大大提高了处理量。这项工作虽已由Frederickson作了介绍,但目前为止尚未见到它正式发表。根据Frederi-ckson的评论,这个方法如果成功的话,那末它很有可能在人类蛋白质组研究中得到应用。
4 结束语
酵母双杂交技术问世不到十年,但已经在蛋白质间的相互作用研究、筛选新的蛋白质、研究蛋白质的功能等诸方面发挥重要作用。双杂交技术固然有其内在的不足之处,如通过双杂交观察到的蛋白质的相互作用在真实情况下不一定发生,也即所谓的假阳性; 假阴性现象也是双杂交分析过程中经常碰到的问题; 一些对细胞致命的蛋白质也不适宜通过双杂交系统来分析。双杂交只是反映蛋白质间能发生作用的可能性,这种可能还必须经过其他实验验证,尤其要与生理功能研究相结合,否则可能会误入歧途。虽然如此,该技术已经成为许多实验室研究蛋白质的相互作用和蛋白质的结构与功能的必要手段。技术的发展与创新、研究成果的迅速共享为建立有机体的全部蛋白质(蛋白质组组分)的相互作用及其功能关系图谱提供了基础和条件。我们不奢望通过双杂交系统能发现所有真实的蛋白质相互作用,但在没有更好的方法问世之前,双杂交技术仍是开展这类研究的有力工具。