科研中的“新星”技术——MicroRNA荧光定量PCR
互联网
Micro RNA简介
微小RNA(microRNA,简称miRNA)是生物体内源长度约为20-23个核苷酸的非编码单链小分子RNA,通过与靶mRNA的互补配对而在转录后水平上对基因的表达进行负调控,导致mRNA的降解或翻译抑制。
是由具有发夹结构的约70-90个碱基大小的单链RNA前体经过Dicer酶加工后生成,不同于siRNA(双链)但是和siRNA密切相关。
第一个被确认的miRNA是在线虫中首次发现的lin-4和let-7,随后多个研究小组在包括 人类、果蝇、植物等多种生物物种中鉴别出数百miRNA。到目前为止,已报道有几千种miRNA存在于动物、植物、真菌等多细胞真核生物中,进化上高度保守。
Micro RNA产生机理
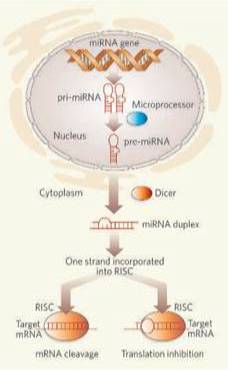
miRNA 是转录后长片段的 RNA 分子(pri-miRNA )的一部分,首先在细胞核内被双链 RNA 特异的核酸酶Drosha处理成70-100 个核苷酸组成的发夹结构 RNA (pre-miRNA )。
这一发夹结构 RNA 运输到细胞质,被另一个双链 RNA 特异的核酸酶 Dicer 剪切,得到 19-23 个核苷酸组成的成熟的 miRNA ,结合在与 RNA 介导的沉默复合物(RISC )类似或者相同的复合物中,这个复合物参与了 RNA 干扰。
由 miRNA 和靶基因 mRNA 的碱基配对引导 RISC降解目的片段或是阻碍其翻译过程miRNA 和它的目的 mRNA 互补的碱基配对决定了这个过程的特异性。通过抑制翻译还是降解发挥作用是由 miRNA 和它的目的mRNA 之间的错配程度决定的,匹配程度高的目的mRNA 将被降解。
由于miRNA 可以对不完全互补的 mRNA 配对来抑制蛋白质的翻译过程,因而每个miRNA 可以有多个靶基因,而几个miRNA可以调节同一个基因。这种复杂的调节网络既可以通过一个miRNA 来经济地调控多个基因的表达,也可以通过几个miRNA 的组合来精细调控某个基因的表达。
对于基于miRNA 调控基因表达的研究的逐步深入,将帮助我们理解高等真核生物的基因组的复杂性和复杂的基因表达调控网络。如下图所示
.jpg)
Micro RNA的特征
已经被鉴定的miRNA据推测大都是由具有发夹结构、约70个碱基大小形成发夹结构的单链RNA前体经过Dicer酶加工后生成的,有5’端磷酸基和3’羟基,大小约21—25nt的小分子RNA片断,定位于RNA前体的3’端或者5’端。
最近3个研究小组分别从线虫、果蝇和Hela细胞中鉴定的100个新miRNA中,有15%跨越线虫、果蝇和哺乳动物基因组具有高度的保守性(只有有1—2 个碱基的区别)。
Lau 和Bartel 实验室的同事更加认为:所有的miRNA可能在其他物种中具有直向同源物(Ortholog,指那些起源于同一祖先,在不同生物体中行使同一功能的基因群就可比作为一个门类,这些类似的基因被称为“直向同源物”)。
Bantam 最早被认为是果蝇中参与细胞增殖的一个基因位点。已知几个包含增强子的转座子插入跨越这个位点的一段12.3kb区域会导致果蝇的眼和翅重复生长,而由转 座子介导的一段跨越该位点的23kb片断缺失则导致突变果蝇个体小于野生型果蝇。
Cohen和同事用一段3.85kb的片断导入21kb片断缺失的果蝇中 使其恢复原来的大小。但是奇怪的是表达这个3.85kb片断中的EST却没有同样的效果。
Cohen将这个片断和疟蚊Anopheles gambiae的同源序列进行比较,发现一段90bp的高度保守区,经过RNA folding program (mfold)发现这个保守序列可以形成发夹结构,使得这个区段很象是一个miRNA的前体。
这个结果经过Northern blot证实突变果蝇的幼体缺少一个21bp的bantam miRNA ,用这个90bp的mRNA前体经过一系列的“功能缺失”—“功能恢复”实验,证实 bantam miRNA在细胞增殖中的作用。
研究人员用计算机程序检索在hid mRNA的3’非编码区找到了bantam的3个潜在的结合位点( hid是果蝇中一个诱导凋亡的基因),并证实 bantam miRNA抑制hid 的翻译而非转录。
miRNA 的表达方式各不相同。部分线虫和果蝇的miRNA在各个发育阶段的全部细胞中都有表达,而其他的miRNA则依据某种更为严谨的位相和时相的表达模式(a more restricted spatial and temporal expression pattern)——在不同组织、不同发育阶段中miRNA的水平有显著差异。
Micro RNA的功能
对microRNA的研究正在不断增加,原因是科学家开始认识到这些普遍存在的小分子在真核基因表达调控中有着广泛的作用。在线虫,果蝇,小鼠和人等物种中 已经发现的数百个miRNA中的多数具有和其他参与调控基因表达的分子一样的特征——在不同组织、不同发育阶段中miRNA的水平有显著差异,这种miRNA表达模式具有分化的位相性和时序性(differential spatial and temporal expression patterns),提示miRNA有可能作为参与调控基因表达的分子,因而具有重要意义。
第一个被确认的miRNA——在线虫中首次发现的lin-4 和let-7,可以通过部分互补结合到目的mRNA靶的3’非编码区(3’UTRs),以一种未知方式诱发蛋白质翻译抑制,进而抑制蛋白质合成,通过调控 一组关键mRNA的翻译从而调控线虫发育进程(reviewed in Pasquinelli 2002)。
bantam miRNA是第一个被发现有原癌基因作用的miRNA。除了lin-4、let-7,已知还有一些miRNA可能参与在细胞分化和组织发育过程中起重要作用的基因的转录后调控,例如mir-14、mir-23 等。
在植物miRNA的研究中有两条线索提示miRNA可能参与植物的发育过程。一是在carpel factory (car) 突变株中3个miRNA的表达水平显著下降。CARPEL FACTORY 是一个类似Dicer的酶,参与植物的发育,其缺失突变株表现为胚胎和叶片发育的缺陷。实验结果提示这种缺陷是由于缺少miRNA加工而造成的。多数的植 物miRNA在某些特定组织中高水平表达也提示他们可能参与了植物组织的发育。
对一部分miRNA的研究分析提示:miRNA参与生命过程中一系列的重要进程,包括早期发育(Reinhart 2000),细胞增殖,细胞凋亡,细胞死亡(Brennecke 2003),脂肪代谢(Xu 2003)和细胞分化(Kawasaki 2003)。此外,一个研究表明,2个miRNA水平的下降和慢性淋巴细胞白血病之间的显著相关,提示miRNA和癌症之间可能有潜在的关系(Calin 2002)。
由于miRNA存在的广泛性和多样性,提示miRNA可能有非常广泛多样的生物功能。尽管对miRNA的研究还处于初级阶段,据推测miRNA在高级真核生 物体内对基因表达的调控作用可能和转录因子一样重要。有一种看法是:miRNA可能代表在一个新发现的层次上的基因表达调控方式。
然而,大多数miRNA的功能仍然是个谜。其机理和功能见下图

Micro RNA识别方法
多个研究小组采用生物化学结合是生物信息学的方法开展对miRNAs的研究工作。由于据推测都是由Dicer酶降解RNA得到的,21—23个碱基大小、有 5’端磷酸基和3’羟基的RNA片断,有的实验室采用改良的定向克隆方法来筛选具有相同特征的小分子——筛选一定大小的RNA分子,连接到3’和5’的适 配子(adapters),逆转录并通过PCR扩增、亚克隆并测序。miRNA前体在基因组上的定位和聚类是通过向基因组数据库查询进行。这个方法有助于 判断miRNAs是否是mRNAs、tRNAs、rRNAs等分子的降解产物。
有的实验室通过一种RNA folding program ’mfold’ 来判断C. elegans 和C. briggsae 之间的高度保守区域是否含有潜在的miRNA前体,然后用Northern Blots的方法来确定这些miRNA是否真的表达了。
尽管有数百个miRNA通过生化或者是生物信息学的方法被鉴别出来,已经鉴别出来的miRNA只不过是沧海一粟,由于很多已经鉴别出来的miRNA是从单个克隆中鉴别出来的,所以可以假设还有很多miRNA在分离和鉴定过程中被“漏掉”了,测序工作还远远不够。
近年来的研究证明,microRNA在干细胞的自我复制、定向分化和组织再生中,也起着十分重要的作用。它是干细胞特性、维持、转化功能的一个关键的调节者,是当前干细胞调节研究的一个重点。最近,Gangara Ju. VK和Lin H在Nature Stem Cell作了一篇综述,系统的总结了microRNA对干细胞调节作用。
Micro RNA与干细胞

但是,microRNA对干细胞的调节作用还远远不止于此。它还参与细胞周期、细胞重建(Reprograming)、多能干细胞生成、信号传递、特异分化、Riches、修复、再生、变异、代谢等所有过程。