肝细胞对疟疾易感性的变化
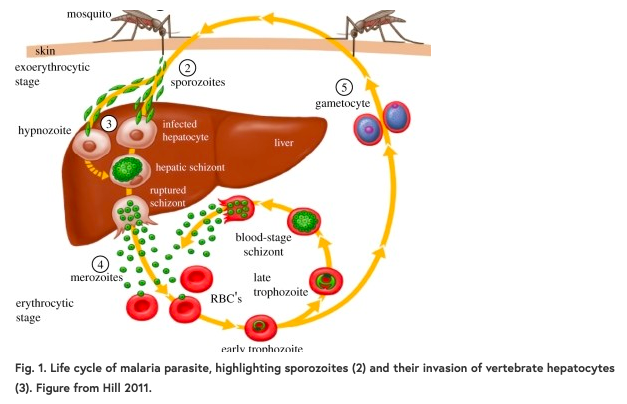
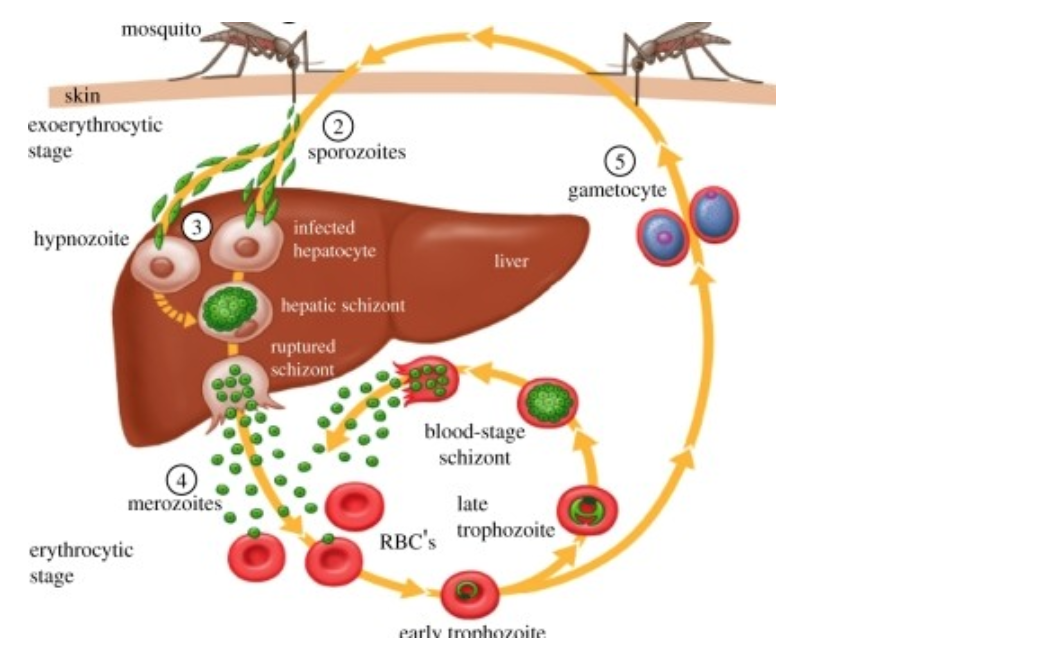
疟疾寄生虫经历了一个复杂的生命周期,每一代在两个生物体中有六个独特的发育阶段。它们在蚊子(媒介)中有性繁殖,在脊椎动物宿主中无性繁殖。在脊椎动物中,当被感染的蚊子在进食时将孢子注入血液,寄生虫就进入宿主体内(图1)。孢子从毛细血管进入宿主肝脏,孢子必须在肝脏中找到并进入肝细胞。这是它们在脊椎动物宿主内的基地,是向前传播的必要步骤。然后这些寄生虫成熟为裂殖体,最终将裂殖子释放到血液中。它开始在红细胞中进行无性繁殖,寄生虫的繁殖和分化进入有性繁殖阶段,最终感染蚊子,再次开始这个循环。
独特的发育阶段为宿主的免疫系统提供了一个不断变化的面具来识别。其中许多已经成为发展的目标(在图1中标注了数字)。每一个阶段都是一个必要的先决条件,最终导致传播,并允许下一代寄生虫。宿主在感染的任何阶段的易感性变化都可能帮助或阻碍寄生虫的发育。例如,缺乏达菲抗原趋化因子受体(DARC)的人不太容易受到间日疟原虫裂殖子的红细胞入侵。这个抗疟的经典例子解释了为什么间日疟原虫在西非没有出现,那里的许多人都是达菲阴性。然而,直到最近我们才知道疟原虫在肝脏初始阶段的感染易感性有任何变化。
Kaushansky和他的同事首先注意到两种实验鼠的肝脏阶段的寄生虫数量的变化。他们进行了第二项研究,以揭示这种变异背后的机制。他们通过多个研究系统发现,ephrin受体EphA2水平较高与疟疾易感性增加有关。首先,他们取EphA2表达不同的小鼠细胞系,将其暴露于啮齿类疟疾约埃利疟原虫的子孢子中,并测量有多少子孢子成功侵入肝细胞,以及1.5小时后有多少细胞被感染。这在活老鼠和人类恶性疟原虫(一种感染人类的物种)细胞系中得到了重复。
在所有的实验中,研究者测量了细胞表面EphA2的表达,比较了EphA2表达的绝对值,并将细胞分为EphA2高表达和低表达两组。无论采用何种方法对EphA2表达进行分类,在所有系统中,EphA2的高表达都意味着宿主易感性的增加(图2A)。这意味着,在表达高水平EphA2的细胞中,感染每个肝细胞的孢子数和感染的总细胞数都更高。随后的实验检查了宿主细胞在较晚时间点的感染情况(寄生虫暴露48小时后),结果表明,在表达高水平EphA2的细胞中,易感性的增加持续存在,并且在较晚的时间点更严重。这意味着EphA2的表达在感染易感性方面可能比早期表现出的作用更大。
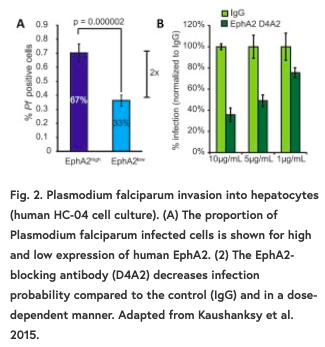
值得注意的是,差异表达与易感性相关,但这并不意味着子孢子数量增加和感染率增加的机制。EphA2是跨膜的,这意味着在肝细胞内和细胞表面存在该蛋白的功能部分。使用小鼠细胞系,研究人员使用抗体来抑制两个区域,一组实验阻断细胞外EphA2和一组实验阻断激酶细胞内区域。当EphA2在细胞外被阻断时,细胞的易感性降低并呈剂量依赖性(Fig2B),而阻断内部结构域时未观察到明显变化。这表明子孢子利用EphA2的胞外部分促进进入宿主细胞。
最令人惊讶的发现之一是这种入侵促进保护了广泛的类群。yoelli疟原虫和恶性疟原虫有很大的不同,这意味着对它们最近的共同祖先的估计是从12米亚(百万年前)到64米亚。宿主,啮齿类动物和人类,也是远亲。寄生虫依赖于一种保守的受体,分布在很大的类群中。这里提供的结果表明,这种易感性的变化在广泛的寄主范围内发现,并被多种疟原虫利用。
肝脏期是疟疾复杂生命周期的一部分,虽然这一阶段不会引起临床症状,但这一阶段感染的减少意味着今后传播受到限制。在这项研究之前,我们对侵入肝细胞的分子基础知之甚少。这项研究确定了肝细胞入侵的一个重要成分,为今后早期预防疟疾感染的研究奠定了基础。