




RNA一度被认为仅仅是DNA和蛋白质之间的“过渡”,但越来越多的证据清楚的表明,RNA在生命的进程中扮演的角色远比我们早前设想的更为重要。RNA干扰(RNA interference)的发现使得人们对RNA调控基因表达的功能有了全新的认识,更因为可以简化/替代基因敲除而成为研究基因功能的有力工具,因此格外引人注意,在2002年度Science评选的10大科学成就中RNAi名列榜首。随着对小分子RNA研究的不断深入 ...
质粒提取的原理http://www.bioon.com 生 物 谷 网 站碱裂解法从大肠杆菌制备质粒,是从事分子生物学研究的实验室每天都要用的常规技术。可是我收研究生十几年了,几乎毫无例外的是我那些给人感觉什么都知道的优秀学生却对碱法质粒抽提的原理知之甚少。追其原因,我想大概是因为《分子克隆》里面只讲实验操作步骤,而没有对原理进行详细的论述。这是导致我的学生误入歧途的主要原因。后来我发现其实是整个中国的相关领域的研究生水平都差不多, ...
质粒DNA的大量提取和纯化 在制作酶谱、测定序列、制备探针等实验中需要高纯度、高浓度的质粒DNA,为此需要大量提取质粒DNA。大量提取的质粒DNA一般需进一步纯化,常用柱层析法和氯化绝梯度离心法。(一)、碱法 1、取培养至对数生长后期的含质粒的细菌培养液400ml,4℃下5000转离心15分钟,弃上清,将离心管倒置使上清液全部流尽。2、称重一般为3-4g. 2、将细菌沉淀物重新悬浮于4ml/1g溶液I中,充分悬浮菌体细胞。(该步骤很 ...

师兄精选:核酸实验板块 牛贴汇总,标题: 【交流】有求必应----DNA克隆URL: http://www.dxy.cn/bbs/post/view?bid=64&id=13864679点击: 4175作者: dnaworm发帖时间: 2009-02-09 15:07:49标题: 【原创】RNA抽提细节系列之四: 解读OD值与RNA纯度的关系URL: http://www.dxy.cn/bbs/post/view?bid=64&id=15495042 ...
PCR反应的最大特点是具有较大扩增能力与极高的灵敏性,但令人头痛的问题是易污 染,极其微量的污染即可造成假阳性的产生.一、污染原因 (一)标本间交叉污染:标本污染主要有收集标本的容器被污染,或标本放置时,由于 密封不严溢于容器外,或容器外粘有标本而造成相互间交叉污染;标本核酸模板在提 取过程中,由于吸样枪污染导致标本间污染;有些微生物标本尤其是病毒可随气溶胶 或形成气溶胶而扩散,导致彼此间的污染. (二)PCR试剂的污染:主要是由于在P ...
(一)酵母双杂交系统的基本原理酵母双杂交系统的建立与发展是基于对真核生物转录调控起始过程的认识。真核生物中存在一种上游激活序列(upstream activating sequence, UAS),其作用是和激活蛋白结合并大大增加启动子的转录速度,从而在转录水平对靶基因表达进行调控。真核细胞转录起始需要反式转录激活因子的参与。很多真核生物的位点特异性转录激活因子是组件式的,通常具有两个可分割开的结构域,即DNA特异性结 ...
最近在做核酸电泳,感觉自己做的差的要死,羞于见人……无奈之下上上网,到处找找资料咯。话说现在做实验似乎已经变成生活的一部分了,不然我怎么感觉这些实验资料每个看起来都这么像人人、豆瓣上分享的生活小贴士……1、让凝胶电泳变得更快,更漂亮方法改进:将电泳电压进行定时变化,例如可以在开始时将电泳电压调节至 100V,大约 15min, 使条带的确可以因为自身片段大小不同而产生较大差别的泳动速度,从而将片段分离,然而现在的分离 或许会间距较 ...
FISH操作前为石蜡包埋组织切片推荐的预处理程序试剂:2% 3-氨基丙基三乙氧基硅烷(3-aminopropyltriethoxysilane, Sigma)、丙酮、二甲苯、酸性亚硫酸钠(sodium bisulfite )、蛋白酶K、2XSSC、0.1M HCl、70%乙醇、85%乙醇、100%乙醇预处理程序:1.将载玻片浸入2% 3-氨基丙基三乙氧基硅烷(3-aminopropyltriethoxysilane, Sigma Chemical Co., St. Lo ...
神奇滤布(Miracloth)上海开米化学有限公司性能参数产品名称: 神奇滤布(Miracloth)英文名称: 神奇滤布(Miracloth)产品货号: 1363488产品规格: 原装试剂级别: 生化级产品产地: 原装品牌商标: 原装供应商: 上海开米化学有限公司价 格: 1200元保存与运输: 干燥保存现货状态: 干燥保存产品更新时间: 2008-6-23 21:19:00详细资料: 实验方法技术资料更多信息:公司展示页面诚信指数:5点浏览人数:46联系我们: 供应商联系卡上海开米化 ...
http://www.uniklinikum-giessen.de/bio/rnai.html这一页提供了许多RNAi公司的链接,值得加分呀!RNAi Drug Discovery CompaniesAlnylam Pharmaceuticals RNAi drug developmentBenitec RNAi stable gene silencing in transgenic miceCenix BioScience RNAi for genome-wide screening ...
在siRNA序列设计时,将潜在的序列用BLAST进行同源序列搜索时,出来的结果好象总是存在和其它编码序列同源的序列,郁闷!是不是可以只看E值?还是只看score?这个问题好菜! 我的目的序列是Human P-glycoprotein (MDR1)中的一个片断,请问出现以下的结果如何判断这个目的序列与其它编码序列不是同源的序列?下面是BLAST的结果: Score ESequences producing significant alignm ...
Version 1.5 of siRNA DNA Designer has been released. The following features have been added.1) BLAST Engine:A BLAST Engine is now included with siRNA DNA Designer 1.5 which permits desktop analysis for potential off-target hits.The Blast engine performs a search for off-target hits using the s ...
1. RNA interference-mediated knockdown of DNA methyltransferase 1 leads to promoter demethylation and gene re-expression in human lung and breast cancer cells.Cancer Res. 2004 May 1;64(9):3137-43.PMID: 15126351 2. A plasmid-based system for expressing small interfering RNA libra ...
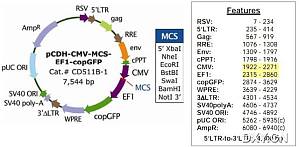
我想把目的基因连接到载体上,有几个问题想请教斑竹和各位战友:1、限制性内切酶的选择:我查看论坛说选择酶切位点需要考虑酶切位点在载体上的距离、酶切温度与连接效率、价格、使用频率等等。最后我综合价格、温度、Buffer选定XbaI和EcoRI,打算购买宝生的酶,相应的Buffer用附带的10×M Buffer。不知道我选择的酶切位点合适不合适呢?2、引物设计:引物结构为:保护碱基+酶切位点+目的基因末端上游引物5’-CCG TCT ...
试验整体方案1.Scrambled (Negative) siRNA Control2.Positive siRNA Control3.Multiple siRNAs to a Single Target4.Monitor Both Target mRNA and Protein Levels5.Use the Lowest Possible siRNA Concentration6.Validation Via Array Analysis and Rescue ExperimentsTo better under ...
Genetic Recombination: Reviews and Protocols形态项:Publisher: Humana PressNumber Of Pages: 272Publication Date: 2004-01-09Sales Rank: 1384602ISBN / ASIN: 1588292363EAN: 9781588292360Binding: Hardcover内容介绍:Genetic recombination is any process in which DNA sequen ...
影响了版面正常操作表示报欠,现将文章部分列在如下:A hypothermic-temperature-sensitive gene silencing by the mammalian RNAiTakashi Kameda, Kenji Ikegami, Yang Liu, Kunihiko Terada, and Toshihiro SugiyamundefinedDepartment of Biochemistry, Akita University School of Medicine 1-1-1 Hondo, Akita 0 ...
很好的常用关于RNAi的网站大全,也许对你有用。Comprehensive List of RNAi Publications (updated daily)RNA info on the WebRNA World Websitehttp://www.imb-jena.de/RNA.htmlRNA interference (RNAi) PDF files on the WebThe Knowledge Foundation: RNA in drug developmenthttp://www.knowledg ...
这个是RNAi对照最权威的文献了,看了以后对对照产品没有疑惑!Whither RNAi? Nature Cell Biology, 5, 489-490 (2003).Whither RNAi?The generally rapid progress of molecular biology methodologies is punctuated on occasion by discoveries that do nothing less than revolutionise the field. Historical exa ...
Nature Protocols 3, - 1452 - 1456 (2008)Touchdown PCR for increased specificity and sensitivity in PCR amplificationDarren J Korbie & John S MattickTouchdown (TD) PCR offers a simple and rapid means to optimize PCRs, increasing specificity, sensitivity and yield, without the need for leng ...

