




多肽质谱仪类型有多种。1、按分析目的可分:实验室多肽质谱仪和工业多肽质谱仪。2、按质量分析器的工作状态可分:静态多肽质谱仪和动态多肽质谱仪。3、按分析规模可分:小型多肽质谱仪和大型多肽质谱仪。4、按联用方式可分:多肽液相色谱质谱联用仪、多肽毛细管电泳质谱联用仪和多肽串联质谱仪等。5、按质量分析器的工作原理可分:四极杆多肽质谱仪、离子阱多肽质谱仪、飞行时间多肽质谱仪和离子回旋共振多肽质谱仪等。
生物制品离心机分类有多种。1、按分离目的可分:实验室生物制品离心机和工业生物制品离心机。2、按结构可分:台式生物制品离心机、落地式生物制品离心机。3、按速度可分:低速生物制品离心机和高速生物制品离心机。4、按转子类型可分:水平转子生物制品离心机和角转子生物制品离心机。5、按分离功能可分:生产型生物制品离心机、制备型生物制品离心机和分析型生物制品离心机。

DAB染色后切片着色呈阴性结果背景:其实免疫组化在DAB染色后一般有四种情况:阳性染色效果很好、阴性染色、非特异性染色很深、阴阳脸(染色不均匀)。而阴性染色是许多初学者经常遇到的头疼问题。我身边有个研究生同学在我们实验室初次涉入免疫组化,听说步骤不难,跟我后面做了几次之后,就自己开始做了,连续做了三次,结果均是阴性染色。很扫兴,不知是什么原因导致的。问题及其解答:免疫组化染色呈阴性结果的原因有哪些?1、抗体浓度和质量 ...
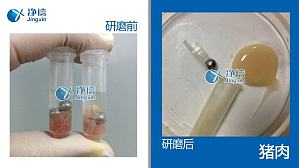
破碎大量样品、低温破碎大量样品、破碎坚硬或韧性较强的样品时,人工手动研磨就成了实验猿的噩梦。当然,也有很多实验猿已经尝试过市面一些研磨机/超声波粉碎机,发现自己那顽固的样品依然无法被完全破碎/破碎过程损坏了目标成分而十分心塞。特别地,一些尤其坚硬/韧性比较强的样品,或要求维持低温条件进行破碎提取的样品,需要结合液氮进行研磨,如:皮肤、硬骨骼、软骨、血管、结缔组织、种子等;使用普通机械研磨方法,很难既保证研磨破碎效果又保 ...
气液填充柱色谱仪的理想载体表面应该无吸附活性和催化活性,在操作条件下不与固定液和样品组分发生反应。但是实际上载体表面完全没有吸附活性和催化活性是不可能的。实验表明,经过灼烧后制成的硅藻土型载体表面既有催化活性,又有吸附活性。当载体表面存在氢键活性作用点时,分离能与硅醇、硅醚形成氢键的化合物(如水、醇和胺等)时,会引起相应组分色谱峰的拖尾。同样,用具有酸性(或碱性)作用点的载体分离碱性(或酸性)化合物时,也会引起相应 ...
核酸质谱仪种类有多种。1、按分析目的可分:化验室核酸质谱仪和工业核酸质谱仪。2、按质量分析器的时空属性可分:时间型核酸质谱仪和空间型核酸质谱仪。3、按结构可分:台式核酸质谱仪和落地式核酸质谱仪。4、按分辨率可分:低分辨核酸质谱仪和高分辨核酸质谱仪。5、按离子化方式可分:电子轰击电离核酸质谱仪、快原子轰击电离核酸质谱仪、电喷雾电离核酸质谱仪和大气压化学电离核酸质谱仪等。
酒精色谱仪分类有多种。1、按分离目的可分:化验室酒精色谱仪和工业酒精色谱仪。2、按功能可分:分析型酒精色谱仪和制备型酒精色谱仪。3、按结构可分:台式酒精色谱仪和落地式酒精色谱仪。4、按操作压力可分:低压酒精色谱仪、中压酒精色谱仪和高压酒精色谱仪。5、按分离规模可分:微型酒精色谱仪、小型酒精色谱仪和大型酒精色谱仪。
气质联用仪种类有多种。1、按分析目的可分:化验室气质联用仪和工业气质联用仪。2、按质量分析器的工作原理可分:四极杆气质联用仪、离子阱气质联用仪、飞行时间气质联用仪和傅里叶变换气质联用仪等。3、按结构可分:台式气质联用仪和落地式气质联用仪。4、按分析规模可分:小型气质联用仪和大型气质联用仪。5、按用途可分:生物气质联用仪、制药气质联用仪、化工气质联用仪、食品气质联用仪、医用气质联用仪、蛋白质气质联用仪、多肽气质联用仪、氨基 ...
色谱仪具有高超的分离能力,分离效果和分离效率远高于蒸馏装置、萃取装置和离心机等其它分离仪器。其主要特点有:1、分离效率高。2、分析速度快。3、灵敏度高。4、样品用量少。5、分离和测定一次完成。6、应用范围广。7、可以和质谱仪等仪器联用。
石油色谱仪类型有多种。1、按分离目的可分:实验室石油色谱仪和工业石油色谱仪。2、按功能可分:分析型石油色谱仪和制备型石油色谱仪。3、按色谱柱形状可分:石油填充柱色谱仪和石油毛细管色谱仪。4、按分离动力学过程可分:石油迎头色谱仪、石油顶替色谱仪和石油洗脱色谱仪。5、按分离规模可分:微型石油色谱仪、小型石油色谱仪和大型石油色谱仪。
气液填充柱色谱仪载体性能的优劣对样品的分离起着重要作用,实际工作中主要依据分离对象、固定液的性质及涂渍量、载体粒度等选择载体。1、分离对象:酸性样品,选用酸洗载体。碱性样品,选用碱性载体。高沸点组分,一般选用玻璃微球载体。强腐蚀性组分,应选用氟载体。2、固定液的性质及涂渍量:当固定液的涂渍量大于 5% 时,可选用白色或红色硅藻土载体。若固定液的涂渍量小于 5% 时,则应选用硅烷化载体。3、载体粒度:常用的载体粒度一般是 80~100 目,为提高柱 ...
液相色谱质谱联用仪类型有多种。1、按分析目的可分:实验室液相色谱质谱联用仪和工业液相色谱质谱联用仪。2、按分析规模可分:小型液相色谱质谱联用仪和大型液相色谱质谱联用仪。3、按质量分析器的时空属性可分:时间型液相色谱质谱联用仪和空间液相色谱质谱联用仪。4、按分辨率可分:低分辨液相色谱质谱联用仪、中分辨液相色谱质谱联用仪和高分辨液相色谱质谱联用仪。5、按离子化方式可分:电子轰击电离液相色谱质谱联用仪、快原子轰击电离液 ...
蛋白质色谱仪分类有多种。1、按分离目的可分:实验室蛋白质色谱仪和工业蛋白质色谱仪。2、按分离模型可分:蛋白质线性色谱仪和蛋白质非线性色谱仪。3、按功能可分:分析型蛋白质色谱仪和制备蛋白质型色谱仪。4、按分离规模可分:微型蛋白质色谱仪、小型蛋白质色谱仪和大型蛋白质色谱仪。5、按操作压力可分:低压蛋白质色谱仪、中压蛋白质色谱仪和高压蛋白质色谱仪。
气液填充柱色谱仪为了取得好的分离效果,特别是分离极性、酸碱性和氢键型化合物时获得对称的色谱峰,对载体常进行酸洗、碱洗、硅烷化和釉化等预处理。一、酸洗:通常用 6mol/L 盐酸浸泡载体,加热处理 20~30 min,然后用水冲洗至中性,用甲醇淋洗、烘干和过筛。也可以用王水或硝酸进行酸洗处理。载体经酸洗后,可除去无机杂质,减小吸附性能,适于分离酸性和脂类化合物。使用时注意,经酸洗的载体表面催化活性较大,不宜分离碱性和醇类化合物。二、碱洗: ...
氨基酸质谱仪分类有多种。1、按分析目的可分:氨基酸化验室质谱仪和氨基酸工业质谱仪。2、按质量分析器的工作原理可分:四极杆氨基酸质谱仪、离子阱氨基酸质谱仪和飞行时间氨基酸质谱仪等。3、按联用方式可分:氨基酸气质联用仪、氨基酸液质联用仪和氨基酸多级质谱仪等。4、按分辨率可分:低分辨氨基酸质谱仪、中分辨氨基酸质谱仪和高分辨氨基酸质谱仪。5、按离子化方式可分:基质辅助激光解吸电离氨基酸质谱仪、电喷雾电离氨基酸质谱仪和大气压 ...
维生素色谱仪种类有多种。1、按分离目的可分:维生素实验室色谱仪和维生素工业色谱仪。2、按结构可分:台式维生素色谱仪和落地式维生素色谱仪。3、按分离规模可分:小型维生素色谱仪和大型维生素色谱仪。4、按流动相物理状态可分:维生素气相色谱仪和维生素液相色谱仪。5、按色谱柱形状可分:维生素填充柱色谱仪、维生素毛细管色谱仪、维生素薄层色谱仪和维生素纸色谱仪。
生命化学是当前化学学科的前沿领域之一,高极性、热不稳定和难挥发的有机高分子化合物难以用气相色谱质谱联用仪进行分离分析。液相色谱仪虽然不受化合物沸点的限制,能对热稳定性差的样品进行分离分析,但定性能力差,应用受到很大限制。这类化合物的分离分析成为化学家面临的重大挑战,开发液相色谱质谱联用仪是迫切需要解决的课题。
落地式质谱仪分类有多种。1、按分析目的可分:落地式实验室质谱仪和落地式工业质谱仪。2、按进样方式可分:落地式探针进样质谱仪和落地式色谱进样质谱仪。3、按分析对象的属性可分:落地式有机质谱仪和落地式无机质谱仪。4、按联用方式可分:落地式气相色谱质谱联用仪、落地式液相色谱质谱联用仪、落地式电感耦合等离子体质谱仪和落地式串联质谱仪等。5、按用途可分:落地式生物质谱仪、落地式制药质谱仪、落地式化工质谱仪、落地式食品质谱仪、落 ...
中华老字号离心机分类有多种。1、按分离目的可分:中华老字号实验室离心机和中华老字号工业离心机。2、按速度可分:中华老字号低速离心机和中华老字号高速离心机。3、按容量可分:小容量中华老字号离心机和大容量中华老字号离心机。4、按分离功能可分:生产型中华老字号离心机、制备型中华老字号离心机和分析型中华老字号离心机。5、按用途可分:中华老字号生物离心机、中华老字号制药离心机、中华老字号化工离心机、中华老字号食品离心机、中华老 ...
高效液相色谱仪分析样品的种类繁多、物理形态广泛、组成及其浓度复杂多变,对分析结果的干扰因素很多,为达到分析目的,样品要进行有效的制备。高效液相色谱仪分析样品的制备包括预处理、提取、净化和浓缩等过程。一、预处理:1、固体样品:含水较低,粉碎过筛。含水量较高,取使用部分烘干后,粉碎过筛。2、液体样品:搅拌混合均匀。3、特殊样品:根据实验要求进行特殊处理。二、提取:1、浸提法(固-液萃取法):将样品浸泡在溶剂中,把固体样品中的某些组分 ...

