




作为第一届四证合一的专业型硕士研究生,面对毕业注定是焦虑、迷茫、挑战、充实。三年即将画上句号,需要奋力去攀爬至人生的新阶梯。总结发现可选择的道路:1. 医院只有

当年阴阳师大火的时候,第一次听身边的朋友说起抽 SSR,心里一惊:难道这玩意儿还能抽?后来明白,此 SSR 非彼 SSR,实验狗说到的 SSR 是这样的:简单重复序列(Simple Sequence Repeat,SSR)也被成为微卫星 DNA,根据其两端互补序列设计引物,通过 PCR 反应扩增微卫星片段,由于核心序列的串联重复数目不同,所以能够用 PCR 的方法扩增出不同长度的 PCR 片段,将这些片段进行凝胶电泳,根据分离片段的的大小确定所研究生物基因的基因型,可以计 ...
其实,在临床上,如果没有什么要紧的事情的话,有些病人还是很好的。有的时候,病人还会主动来夸你,夸的人小心肝一颤一颤的~(哈哈哈哈哈哈,美滋滋一整天……)今天早上查房的时候,被一个阿婆夸长得俊,莫名开心哈哈哈哈大家都有被病人夸过吧~大家都被病人夸过什么?(长的俊?技术好?(来自丁香园站友@熊欣欣)@mazeee:一个师兄值夜班的时候,接诊了一位带着孩子来看病的年轻妈妈,师兄面相成熟又带了口罩,看完以后,年轻妈妈十分感激,拉着孩 ...
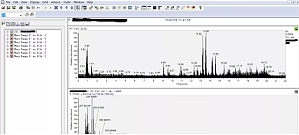
代谢组学是现在医药领域用来探究某种疾病或药物作用机制的一种热门手段。目前代谢组学的研究主要采用三种技术平台:LC-MS、NMR 和 GC-MS。这三种技术各有利弊,相对来说,LC-MS 对于代谢物的检测最为全面。对于 LC-MS 代谢组学方法得到的代谢物指认,Xcalibur 软件和 Metlin 数据库最为实用。下面我们就讲一下 Xcalibur 软件和 Metlin 数据库的使用方法:1 .找到想要解析的数据,双击,出现如下界面,激活右上角色谱图光标,使其变绿, ...
在近日召开的 「第三届中国医疗卫生行业 EAP 论坛」上,专家亮出一组数据:根据去年国内医生群体生存调研显示,77% 的医生一周工作超过 50 小时,43.5% 以上表示每月最长留宿 8 次以上 ,69.4% 的正高级医生每月参加会议 4 次以上。(丁香园战友 @古小医)@爬墙头等红杏:休假是不可能休假的,这辈子都不可能休假的,不加班就是好的!!!@xiaolong1a:还没休过年假。医院说休年假不给算全勤。@殇槿:有是有,没有休过,科室人不够,休不了,天天加班到晚上 8.9 点,以前 3 天一夜班 ...
如题,欢迎大家畅谈您的丁香园缘起。(来自丁香园站友@大雪纷飞蓝天白云)@wj7994712 我是在其论坛被介绍过来的。。。@ymg2000 我是找资料时被吸过来的@死亡呼吸 我是同学介绍来的,等到我的 id 被他们强行共享时才发现上当了,原来他们的 id 都还是 0 分。@wxdong100 我是听别的医生介绍的。可是当时访问量并不大, 内容也并不多, 可现在却大不一样了, 04 年 4 月份我一来这, 就被这吸引住了, 现在几乎天天都来.@GavinDu 同学介绍来的@suncat 在另一个医学生 ...
我 14 年毕业的本科生,毕业后找了个社区医院,同年就被送出去规培了,全科规培,现在结束了,会自己医院上班,干妇产科的,可是规培三年妇产科只安排了一个月,现在什么都不会,又被送到二级的县医院进修,才发现他们医院有好多本科生,和我同时上班的,现在都能单独值班、处理突发情况了,我连皮毛都不懂,差距太大了!想跳槽往二级医院,可是三年已经浪费了,而且规培证书都是全科方向的,往上跳的机会太小了,想来想去只有考研了…… 可是如果考研走了,我要 ...

不知道师兄师姐是不是跟你说过综述都是大牛写的,像我们这样的研究生吭哧吭哧做实验就好了。对于大部分学校而言,发表一篇 SCI 论文,硕士就能达到小论文的要求了,博士综述型文章也可以算一篇。若 N 为毕业要求 SCI 论文数目,那么一篇做实验一边写综述的小明同学就只要 N-1 篇研究性文章了。不过大家都说综述不是我们普通人能发的,那么平民研究生小明同学到底能不能发出文章呀。诚然一些顶级综述都是邀稿型,以国家某重点实验室菜鸟小硕为例,能接收的非邀 ...

还记得 2018 年年初,学霸平台曾推出一篇《狗年说狗 | 那只改变美国科研界的宠物狗》时光荏苒,不知不觉,2018 年已经过去一大半,你还记得年初制定的小目标吗?在此,学霸君特意推出狗年说狗的姊妹篇--那些植物中的「狗狗」,回想年初的誓言,让我们抓紧狗年的最后一个季度去实现 2018 年的梦想。听说集齐动物、植物的「狗狗」们,可以在生物学霸里召唤真龙吆,让我们一起来看看植物中都有哪些好玩的「狗狗」吧。NO.1-- 狗尾巴草说到植物中的「狗狗」,有一种特别 ...
2018 年起,生物学霸联合丁香人才推出招聘服务,我们不仅解决大家科研上的问题,我们还想办法解决大家的就业问题,是不是挺贴心的呢,记得给我们点赞鼓励哦。今天给大家推出的是中科院生化与细胞研究所博士后招聘。中科院生化与细胞研究所中国科学院生物化学与细胞生物学研究所(简称:生化与细胞所)成立于 2000年,是中国生命科学领域的一个旗舰研究所,前身是原中国科学院上海生物化学研究所与原中国科学院上海细胞生物学研究所,历史可追 ...
成为一名医生,护士,是一种什么感受?你还记得成为医生走向工作岗位的第一年如何?8月19日 中国医师节之际,我们征集到百名戳心回答:我想对第一年工作的自己说:@ 调皮鬼:我的第一份工作是急诊科医生,在我工作的第二年考上了本院的编制,做了一名心内科医生。我想对第一年工作的人说:如果当年更努力一些,也许结局会更好一些,现在努力还不晚,多向前辈请教,遇到不懂得就问,多干多练,终有一天你可以独当一面。@ 晚秋:儿科医生。执业医师证是 06 年 12 月考出 ...
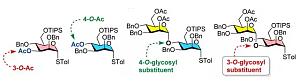
作为一名资深的化学研究生,利用 chemdraw 软件来画化学结构式与反应式已经是得心应手,给原子编号和化学键上色等一些美化修饰手段是驾轻就熟,对于研究糖化学的工作者来说,画好的一连串的椅式构型的糖分子犹如一幅优美的画卷。但是也会看到有的文献图中糖的化学式中不同糖环的内部颜色还不同,相比于不同化学键的不同颜色来说显得更直观清晰,也显得更高大上,正如图中所示。本人第一次看到的时候也是惊叹不已,于是就在 chemdraw 软件 ...
①神经科学家逆转了“天才/疯子”的一些行为症状;②“一夜好眠”即将来临;③Cell:人体蛋白质个体差异12%是由性别和饮食决定的;④什么物质能阻止神经细胞退化?;⑤《Nature》:Cryo-EM技术完成不可能完成的任务
现如今,人口老龄化问题十分严峻,医生行业也是如此。大多数患者的「传统思想」都认为,嘴上无毛,办事不牢。医生越老越吃香,尤其是中医,越老越有人找。年轻人刚入行挣得也少,又导致学医的年轻人改行的越来越多。大家如何看待医生老龄化现象?(来自丁香园站友@thankyouu)@小东邪黄药师: 我觉得主要是医疗环境和薪酬,压力对比,还有就是社会对医疗的高要求,低投入,学习哭累,工作死板不够灵活,导致吸引力下降。@gimma_nuts: 还有些人 ...
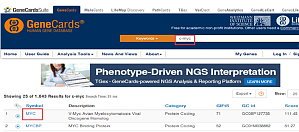
如果要验证某一个基因是否有形成 G4 联体的可能性,首先需要在其序列上找到富含 G 序列的片段,而最有可能形成 G4 联体的区域包括启动子区、5' 非编码区,所以我们的查找范围就锁定在以上两个区域。找到所需序列之后再借助工具分析找到富含 G 序列的片段。1首先在 genecards 上搜索你需要的基因,这里以 MYC 基因为例。点击查找到的第一个基因。2点进去就是这个页面啦,找到基因的编码链序列,就是大家都知道的 NC 号了。3然后跳转到 NCBI 的页面,接下来就要去找我们需 ...
什么?我们也能有假期?可能大部分同道就算有公休年假的也不敢修,比如我... 怕被领导同事惦记...像春节、国庆这种稍微法定小长假,还要值班。但还是能有空闲的时间的是吧~ 来说说你的安排呀~我选择补觉(来自丁香园站友 @ Doctor 先生_)@美食家汉尼拔医生:16 年国庆跟同事调了班,7 天去了贡嘎重装徒步,平时打系统内的足球赛,每周跟科里打排球,每周按计划表撸铁四次,回家滑雪,3 月份打算去涛岛潜水靠 ow,或者尼泊尔走 act 线。是不是不像医生做的事情。@天生 ...
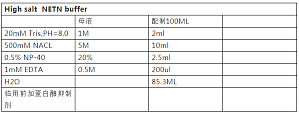
近年来关于组蛋白的研究越来越多,组蛋白各种各样的修饰有各种各样的功能,想要做跟组蛋白相关的研究,组蛋白提取是一个必不可少的技能了。你还在为组蛋白如何简单方便的提取而烦恼么,来,让学霸君教你一招。组蛋白有四种类型:H2A、H2B、H3、H4,组蛋白是染色体基本结构蛋白,因富含碱性氨基酸 Arg 和 Lys 而呈碱性,可与酸性的 DNA 紧密结合。因为组蛋白是偏碱性的,因此我们可以使用酸法来提取。下面学霸君就仔细介绍一下实验操作步骤啦:1、将 3- ...

看到一个有趣的话题,医生到底是不是一个服务行业?有人说:在《中华人民共和国职业分类大典》,医生是技术人员。但同时,医生也属于公共服务业。有人说:「现在在中国,如果医生不把自己归于服务业还能活下去吗?」,「态度不好就会被砍啊……」所以,在你们看来(心目中),医生是服务业吗?(来自丁香园站友@用户名已占用 555)@gchend: 服务业就会构成绝对的消费模式!即病人消费就是上帝,你就得听他的按他说的做!所以国家的定位不清,无法保证医疗技术 ...

DAVID(the Database for Annotation, Visualization and Integrated Discovery),它是一个生物信息数据库,整合了生物学数据和分析工具,为大规模的基因或蛋白列表(成千上百个基因 ID 或者蛋白 ID 列表)提供系统综合的生物功能注释信息,帮助用户从中提取生物学信息(想了解更多,转战度娘~)。Tips: 通过 DAVID 对肿瘤相关基因做功能注释和富集分析。打开 DAVID 平台http://david.abcc.n ...

以前,医生一直是人们眼中的 「高大上」职业,但随着 「收红包」、「大处方」等问题的曝光,医生的形象大打折扣,有的地方甚至出现了严重的医患冲突。如今的医生,到底是怎样一个群体?你眼中的医生是什么样子?在大家学医之前,大家眼中的医生是什么样的呢?学医前:高大上、收入高,人上人?学医后:苦穷丑、累死狗…@傻某二:我错了我错了我错了……以前一直以为医生就是每天坐在办公室喝喝茶收收红包炒鸡爽的,毕竟学医辣么辛苦,工作了过得舒服也是应该的。于是 ...

