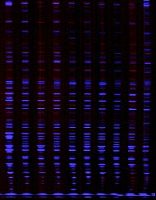
AFLP: not only for fingerprinting, but for positional cloning
互联网
<center> <p><font></font> </p> <p><font></font> </p> <h2> <center><font>Mannie Liscum(1)</font> <font> and Paul Oeller(2)</font> </center> </h2> <p><font></font> </p> <h3> <center><font>Department of Plant Biology</font> </center> <center><font>Carnegie Institution of Washington</font> </center> <center><font>Stanford, CA 94305</font> </center> </h3> <p><br /> <br /> <br /> <b><font>(1)Current address:</font> </b> <i><font>http://www.biosci.missouri.edu/liscum/LiscumLabPage.html</font> </i> <font> (site contains updated information)<br /> Division of Biological Sciences<br /> 105 Tucker Hall<br /> University of Missouri<br /> Columbia, MO 65211<br /> phone: 573-882-2672<br /> fax: 573-882-0123<br /> email: mliscum@biosci.mbp.missouri.edu </font> </p> <p><font></font> </p> <p><font></font> </p> <p><font><font><b>(2)Current address:</b> DNA Plant Technology Corporation<br /> 6701 San Pablo Avenue<br /> Oakland, CA 94609<br /> phone: 510-547-2395<br /> fax: 510-547-2817<br /> email: oeller@dnap.com</font> </font> </p> </center>
NOTE: The protocol presented below is based on the Amplified Fragment Length Polymorphism (AFLP) technology developed by Marc Zabeau and colleagues at Keygene N.V., Agrobusiness Park 90, P.O. Box 216, NL-6700 AE Wageningen, Netherlands (Zabeau, 1992; Zabeau and Vos, 1993; Vos et al., 1995). The AFLP technology is covered by patents and patent applications owned by Keygene N.V. Both, Life Technologies (Gathersberg, MD, USA) and Perkin Elmer (Applied Biosystems Division, Foster City, CA, USA) market research kits (under license) for AFLP fingerprinting of plant DNAs.
Background Rational
AFLP as developed by Keygene, was designed as a highly sensitive method for DNA fingerprinting to be used in a variety of fields, including plant and animal breeding, medical diagnostics, forensic analysis and microbial typing, to name a few. We are using this technology to generate DNA based markers for cloning genes involved in phototropic responses in higher plants that have only been identified genetically by mutant phenotype(s) (see Liscum and Briggs, 1995). We have had tremendous success to date using the technology decribed below (see pg. 4). (Recently, Thomas et al. (1995) reported the use of AFLP technology in the identification of tightly linked markers flanking (within 15.5 Kb) the Cf-9 resistance gene of tomato. Although the Cf-9 gene had been previously isolated via transposon tagging the rapid identification of markers within 15.5 Kb of the locus provide support for the contention that AFLP technology can be exploited for gene isolation by positional cloning.) The "guts" of how AFLP works is summarized in the following paragraphs.
The power of AFLP is based upon the molecular genetic variations that exist between closely related species, varieties or cultivars. These variations in DNA sequence are exploited by the AFLP technology such that "fingerprints" of particular genotypes can be routinely generated. These "fingerprints" are simply RFLPs visualized by selective PCR amplification of DNA restriction fragments. In order to give a brief summary of the working theory behind AFLP we will describe the technique as we use it to generate AFLPs between two Arabidopsis ecotypes, Columbia and Landsberg erecta. 1) Genomic DNA from each ecotype is digested to completion with two restriction enzymes, one EcoRI (having a 6 bp recognition site) and the other MseI (having a 4 bp recognition site). EcoRI cuts ~ every 2-2.5 Kb in Arabidopsis, while MseI cuts ~ every 300-400 bp. Thus a large number of fragments are generated with relative abundances as such: MseI-MseI >>> MseI-EcoRI >>> EcoRI-EcoRI. As discussed below we target the MseI-EcoRI and EcoRI-EcoRI fragments for analysis. 2) Specific ds oligonucleotide adapters (~ 25-30 bp) are ligated to the restricted DNA fragments. 3) Oligos homologous to the adapters, but having extensions at the 3'-end are used to amplify a subset of the DNA fragments. These extensions can vary in length from 1 to 3 bp, but are of defined length for a given primer. The sequence of the extension can also vary from one primer to another but is of a single, defined sequence within a given primer. The selective nature of AFLP-PCR is based on the 3' extensions on the oligonucleotide primers. Since these extensions are not homologous to adapter sequence, only plant DNA fragments complementary to the extensions will be amplified due to the inability of Taq DNA polymerase, unlike some other DNA polymerases, to extend DNAs if mismatches occur at the 3'-end of a molecule that is being synthesized. Therefore only a subset of the entire genome is amplified in any reaction. For example, if 2 bp extensions are used only one in 256 molecules is amplified assuming Taq polymerase cannot tolerate mismatches as discussed above. However, since MseI-MseI fragments predominate the population of fragments to be amplified, we need to further limit the number of fragments that are actually visualized so that a manageable number is observed. We do this by labeling the primer directed against the EcoRI-adapter sequence since MseI-EcoRI and EcoRI-EcoRI fragments will be a more limiting subset of the total DNA fragments. Finally, the amplified DNAs are separated on a polyacrylamide gel (sequencing type) and an autoradiograph is generated.
By labeling the EcoRI-directed primer and using 2 bp extensions on both primers we typically observed 100-200 bands on the autorad from any given primer pair. Only a subset of these total bands are polymorphic between two related individuals, such as Arabidopsis Columbia and Landsberg erecta ecotypes. Any given AFLP primer pair generates on average 10-20 ecotype specific bands (conservatively), thus given 256 possible primer pair combinations when using 2 bp selective 3' ends, 2500-5000 markers can be generated quite rapidly. Given a genome size of 100 Mbp for Arabidopsis, and assuming equal recombination frequencies over the entire genome (or equal distribution of markers throughout the genome), generation of 2500 markers would place any pair of markers within 40 Kb of each other, on average. This level of physical linkage would result is the generation of markers that are _ 0.3 cM (0.3% recombination) within the mutant locus of interest. Thus AFLP should eliminate the "walk" typically associated with non-T-DNA-tagged mutants, and substitute the "hop" or "land".
AFLP Protocol (Abridged Version 1.3, 12/95)
1.0 Generate polymorphic recombinant F2 (or F3) population
Outcross desired mutant in one genetic background (parental ecotype of the mutant hence referred to as WT1) to a wild-type plant of another genetic background (WT2) known to be molecular genetically polymorphic to WT1. [We have used three WT ecotypes of Arabidopsis for our work (Columbia, Landsberg erecta, & WS) and each is about equally polymorphic from any other. There is no reason to believe that other ecotypes would not be equally suitable.] Self the resultant F1 seed and select homozygous mutants from the subsequent F2 generation. Collect tissue from homozygous mutant F2 plants, as well as WT1 and WT2 plants, for DNA isolation as described below in Sec. 1.1 . Self homozygous mutant F2s and collect seed for future use. It is also helpful to save a few non-mutant F2s for tissue collection and F3 seed collection as they may be useful controls for future experiments.
1.1 Isolate genomic DNA
NOTE: Use any mini(micro)prep that yields good quality DNA. CsCl-purified DNA is not necessary. The main contaminants of concern are carbohydrates. If the A260/A230 is _ 2.2 the nucleic acid should be of adequate quality. Dark adapting the plants for 2-3 d prior to tissue harvest is usually enough to insure low carbohydrate content.
Microprep1.) Dark adapt 3-4 week old plants (just beginning to bolt) for 2-3 d, then harvest tissue (1-4 rosette leaves) into 1.5 mL microfuge tubes w/ liquid N2. Grind tissue on liquid N2 with pellet pestle (Kontes Scientific Glassware/Instruments; cat #749520-9001 for pestles only, #749520-0000 for tubes and pestles). [Using an electric drill to hold and rotate the pestle works extremely well!] Hold ground tissue at _ -80° C until extraction.
2.) For extraction of nucleic acids, add 750 µL extraction buffer (see below) to ground tissue, vortex to mix thoroughly, and incubate @ 65° C for 10 min.
Extraction Buffer
50 mM Tris, pH 8.0
10 mM EDTA, pH 8.0
100 mM NaCl
1.0 % (w/v) SDS
10 mM ß-mercaptoethanol (add just before use)3) Add 150 µL 5 M K-acetate, vortex, incubate on ice for 20 min. Spin @ 12K x g in microfuge for 10 min.
4) Transfer 750-800 µL supernatant to new microfuge tube and add an equal vol of isopropanol. Immediately spin down nucleic acids by centrifugation @ 12K x g for 1 min. [Some small pieces of tissue may pellet at this stage, but they will be removed in the next step.]
5) Wash pellet w/ 75% EtOH and resuspend in 200 µL H2O. Spin @ 12K x g , 2 min. Remove 180 µL of supernatant to new tube and add 20 µL 3 M Na-acetate (or K-acetate). Mix and add 500 µL EtOH, then incubate @ -20°C for _ 10 min. Pellet nucleic acids by centrifugation (12K x g, 5 mn). [This step will remove chlorophyll as well as any debris carried over from step 4.]
6) Wash pellet w/ 75% EtOH, dry and resuspend in 10-50 µL TE (pH 7.5).
7) Quantitate nucleic acids and assess purity (A260/A230; A260/A280). [This microprep yields nucleic acids with A260/A230 > 2.0 and A260/A280 ~ 2.0. Although most of the nucleic acid is probably RNA, in practice there appears to be a sufficient amount of DNA for reproducible AFLP results if one follows the protocols below.]
1.2 Restriction of DNA
(Recipe is for one sample)0.5 µg genomic DNA (total nucleic acid from mini(micro)prep)
5 U EcoRI
5 U MseI
8.0 µL 5x-Pharmacia "One-Phor-All+" buffer [10x = 100 mM Tris-acetate, pH 7.5, 100 mM Mg-acetate, 500 mM K-acetate) w/ 250 ng/µg BSA (=5x OPA+- BSA)
Q.S. to 40 µL w/ dH20- incubate @ 37° C for ~ 3 hrs
[Note: The digestion should not be done for significantly longer than 3 hrs, as it is necessary to have active enzymes present during the ligation step (Sec. 1.3 ) to ensure complete digestion and ligation]
1.3 Ligation of adapters
Adapter PreparationEcoRI-adapter = 5'-CTCGTAGACTGCGTACC (EcoRI-oligo.1) CTGACGCATGGTTAA-5' (EcoRI-oligo.2)- mix 1.7 µg EcoRI-oligo.1 and 1.5 µg EcoRI-oligo.2, 3 µl 10X One-Phor-All+,Q.S. to 60 µL w/ dH2O, heat to 95° C, and allow to cool to RT slowly. This gives a final concentration of 5 pmoles/µL and makes enough adapter for 60 ligations (see below).
MseI-adapter = 5'-GACGATGAGTCCTGAG (MseI-oligo.1) TACTCAGGACTCAT-5' (MseI-oligo.2)- mix 16 µg MseI-oligo.1 and 14 µg MseI-oligo.2, 3 µl 10X One-Phor-All+,Q.S. to 60 µL w/ dH2O, heat to 95° C, and allow to cool to RT slowly. This gives a final concentration of 50 pmoles/µL and makes enough adapter for 60 ligations.
[Note: Adapter oligos should not be phosphylated, this prevents adater self ligation. Both adapters are engineered such that the ligation "kills" the restriction site to which the adapter is ligated.]
Ligation RxnAdd 10 µL of following mix to each 40 µL digestion rxn:
(recipe is for one ligation rxn)
1.0 µL EcoRI-adapter
1.0 µL MseI-adapter
1.0 µL 10 mM ATP
4.0 µL 5x-OPA+-BSA
1 U T4 ligase (Pharmacia)
Q.S. to 10 µL w/ dH20
- Incubate @ 37° C for 3 hrs to O/N
2.0 Pre-amplification of template DNA
[Note 1: This pre-amplification step helps to "clean up" some background noise that is observed on autorads of gels where non-preamplified DNA was used as a template. Furthermore, for every 1 µL of total ligated nucleic acid that is preamplified one will have 150 µL of template for further amplification (see below).]
[Note 2: For this and subsequent PCR we use an MJ Research thermocycler (# PTC100; Watertown, MA, 1-800-729-2165). This thermocycler accepts 96-well microtiter plates, which is a more efficient way to amplify the multiple samples required for AFLP.]
[Note 3: The AFLP procedure reported in NAR by Vos et al. (1995) uses a preamplification with both EcoRI and MseI primers having 1-bp, 3'-extensions, followed by AFLP-PCR with primers having 3-bp, 3'-extensions, whereas our preamplification uses only our "EcoRI-oligo.1" (adapter oligo with no extension) (see below), and our AFLP-PCR uses only 2-bp, 3'-extensions (see 2.2). Life Technology and Perkin Elmer protocols use the Vos et al. (1995) preamplification and AFLP-PCR. In our experience the single primer amplification (EcoRI-oligo.1) works quite well in comparison to the 1-bp two primer preamplification without reducing the number of polymorphic bands in the subsequent AFLP-PCR reactions. However, it should be noted that there is alot of room for "personalizing" the preamplification and AFLP-PCR steps to fit ones needs.]
Pre-amplification cocktail (recipe is for one rxn)0.5 µL EcoRI-oligo.1 (@ 50 ng/µL)
0.8 µL 5 mM dNTPs
2.0 µL 10x-Promega Taq buffer [500 mM KCL, 100 mM Tris-HCl, pH 9.0 (@ 25° C), 1.0% Triton X-100; Mg-free)
1.2 µL 25 mM MgCl2
0.08 µL (0.4 U) Taq Pol (Promega)
14.42 µL dH2O
PCR reactions (recipe is per rxn)
1 µL cut/ligated nucleic acid (from above) [equals 10 ng total nucleic acid)
19 µL pre-amplification cocktail- amplify using "PRE-AFLP" as listed below:
"PRE-AFLP" thermocycle profile
1) 94° C, 2 min
2) 94° C, 30 seci
3) 50° C, 30 sec
4) 72° C, 1 min
5) repeat steps 2 to 4, 34 times
6) hold @ 4° C
- Do NOT perform a "hot start".
-dilute PCR products 1:10 w/ TE (pH 7.5) and store @ 4° C (or -20° C for long term) -
2.1 Primer labeling (w/ 33P or 32P)
(recipe is for a 50 µL labeling rxn)§10.0 µL EcoRI-primer† (@ 50 ng/µL)
10.0 µL g33P-ATP (or g32P-ATP) [3000 Ci/mmol]
5.0 µL 10x-OPA+ (no BSA)
1.0 µL (5-10 U/µL) T4 polynucleotide kinase (PNK)
24.0 µL dH2O
§50 µL labeling rxn makes enough "hot" primer for 100 PCR rxns.
† The basic EcoRI-primer is:
5'-AGA CTG CGT ACC AAT TCx yz-3'
where, x, y, and z represent the selective bases on the 3' end of the oligo.[We typically use only 2 bp selective extensions (x and y, no z in our primers). x, y, and z are specific and constant bases within a single primer but vary from primer to primer. Thus with 2 bp selective extensions one will have 16 independent primers.]
2.2 AFLP-PCR
AFLP cocktail (recipe is for one rxn)0.5 µL labeled EcoRI-primer (@ 10 ng/µL)
0.6 µL unlabeled MseI-primer‡ (@ 50 ng/µL)
0.8 µL 5 mM dNTPs
2.0 µL 10x-Promega Taq buffer (Mg-free)
1.2 µL 25 mM MgCl2
0.08 µL (0.4 U) Taq Pol (Promega)
13.82 µL dH20
‡ The basic MseI-primer is:
5'-GAT GAG TCC TGA GTA Axy z-3'like with the EcoRI-primer, x, y, and z represent selective bases that are specific and constant within a primer but vary between primers. [We typically use 2 bp selective extensions in our MseI-primers. The Zabeau group, as well as commercially available kits use 3-bp selective extensions in combination with 1- bp selective preamplification (see 2.0, Note 3) ]
PCR reactions (recipe is for one rxn)
1 µL preamplified DNAi
19 µL PCR cocktail- amplify using "AFLP" as listed below:
"AFLP" thermocycle profile
1) 94° C, 2 min
2) 94° C, 30 sec
3) 65° C, 30 sec, -0.7° C per cycle starting next cycle
4) 72° C, 1 min
5) repeat steps 2 to 4, 11 times
6) 94° C, 30 sec
7) 56° C, 30 sec
8) 72° C, 1 min
9) repeat steps 6 to 8, 25 times
10) 72° C, 2 min
11) hold @ 4° C
3.0 PAGE of PCR products
Sample preparation§ can use 0.5x TBE as well but 1x TBE often gives clearer banding.1) Mix each rxn w/ an equal vol of 2x formamide buffer (98% formamide, 10 mM EDTA, pH 8.0, plus bromophenol blue & xylene cyanol for tracking dye).
2) Incubate samples @ 90° C for ~ 45 min [this reduces the vol and thus increases the specific activity of the samples].
Electrophoresis
1) Load sample on gel [5.0% acrylamide/bisacrylamide (19:1), 7.5 M urea, 1x TBE§ and run @ 40-50 W (constant wattage) [40-50 V/cm, like for sequencing gels] with 1x TBE as the running buffer.
2) Run samples until the xylene cyanol is about 2-3 cm from the bottom of the gel and stop. [When 1x TBE is used in the gel and running buffer, and the gel is electrophoresed @ 50 W, it takes about 3.5-4 h for the xylene cyanol to get to the indicated position.]Fixing & autoradiography
1) Fix gel in 10% acetic acid for 30 min.
2) Dry gel onto filter paper (E&K Scientific Products, cat# 2388-4567, 46 cm x 57 cm sheets) for _ 1 hr w/ heat.
3) Generate autoradiograph using Kodak BioMax film (or like product from another supplier). [This film gives nice O/N exposures when using either 33P or 32P as the label.]
4.0 Linkage analysis
Analyze F2 (or F3) recombinants relative to the two parent ecotypes (WT1, the genetic background of the mutant; and WT2, the polymorphic wild-type to which mutants were outcrossed, see Sec. 1.0) used as the backgrounds to generate the recombinants. [We typically use Columbia as the parental stocks (WT1 line) for mutagenesis and Landsberg erecta as our WT2 line.] Linkage is assessed by finding polymorphic bands specific to the WT2 parent that are missing in a large sample of F2 individuals (see below for suggested population size). The reason for looking for the absence of a WT2-specific band rather than the presence of a WT1 band is the inability to discern heterozygotes from homozygotes. For example, any F2 individual having a WT1-specific band could be either linked at one chromosome or both. The absence of a WT2-specific band requires therefore that the DNA specified by that polymorphic marker is homozygous for WT1 DNA in the F2 recombinant; or both chromosomes contain WT1 DNA that is linked to the locus of interest. The pitfall of this approach is that the presence of a WT2-specific band in an F2 individual can therefore be either WT2/WT1 or WT2/WT2. For example, if 1 in 100 F2 individuals has a WT2-specific band of interest the recombination percentage (linkage) can be either 0.5% (1 chromosome in 200) or 1% (2 chromosomes in 200). We therefore suggest examining at least 400-500 chromosomes to have high confidence in the linkage results. Any WT2-specific bands of interest are then subcloned and used to generate linked DNA fragments and oligos for library screening as described below.
4.1 Band isolation and subcloning
Band isolation1) Perform a standard AFLP-PCR rxn with the WT1 and WT2 parents using the primer pair(s) (EcoRI-primer should be end labeled) that result in an excluded WT2 band(s) in the F2s
2) Precipitate the PCR products. Adjust the rxn products to 0.3 M Na-acetate and add 2.5 vol EtOH, incubate @ -20° C for _ 20 min, then spin @ 12K x g for 5 min. Resuspend each DNA in just enough 2x formamide buffer to load into 1 lane on a standard PA-AFLP gel. Run gel, dry (DO NOT FIX), and generate an autorad as per normal (see Sec. 3.0).
3) Cut out band of interest from dried gel (using autorad as the guide) and isolate the DNA via the "crush & soak" method of Sambrook et al. (Vol 1, pp. 6.46-6.48). Precipitate the DNA and resuspend in 10 µL of TE (pH 7.5). Use this DNA as the template for PCR amplification with "Universal AFLP Cloning"-primers (U-EcoRI and U-MseI) as described below.
PCR for subcloning
- Set up PCR rxns as per Sec. 2.0 except that the PCR cocktail contains 0.5 µL of each U-EcoRI- and U-MseI-oligos£ and only 13.92 µL dH2O instead of 14.42 µL per rxn. Use 1 µL of the DNA in step 3 above per rxn. Amplify with "PRE-AFLP".
£ U-EcoRI- and U-MseI-oligos are as follows:
U-EcoRI = 5'-GCG GAA TTC CGT AGA CTG CGT ACC AAT TC-3'
- this oligo has an EcoRI site in its 5' end -
U-MseI = 5'-GCG CTG CAG GAC GAT GAG TCC TGA GTA A-3'
- this oligo has a PstI site in its 5' end -
Subcloning4.2 Verification of cloned markers1) Digest a desired amount of above PCR product with EcoRI and PstI, and ligate into like restricted plasmid vector of choice. [NOTE: There is some probability that the fragment of interest contains a PstI site. Cloner beware!]
2) Transform E. coli with recombinant plasmid and select for plasmid+ colonies by standard methods.
[NOTE: We typically use a two step verification of marker clones (as described below); one based on PCR and one based on Southern blot analysis.]
Clone verification I: Colony PCR[NOTE: This amplification results in a single stranded probe that is complementary to the WT2-specific band observed on a standard AFLP gel (see linkage analysis, Sec. 4.0) .]1) Pick plasmid+-selected colonies w/ toothpick
2) Dot onto new LB plate plus selective antibiotic
3) Inoculate 10-25 µL of digestion buffer (see below) with remaining cells on toothpick
Digestion Buffer4) Incubate @ 55° C, 2 hrs
8.9 vol TE, pH 8.0
1.0 vol 10% (w/v) Laureth-12
0.1 vol 20 mg/mL Proteinase K5) Heat inactivate Proteinase K @ 95° C, 10 min
6) Use 1 µL of digestion for PCR
7) Amplify as per AFLP-PCR (Sec. 2.2) using the EcoRI- (end-labeled) and MseI-primers that were used in step 1 of Sec. 4.1 (Do not pre-amplify! Just use DNA from digestion directly.). Adjust the volume of PCR cocktail such that the total rxn (cocktail + template) is 20 µL. Also amplify both WT1 and WT2 parents and any recombinants that have the WT2-specific band.
8) Compare banding patterns of amplified E. coli DNA (plasmids) with that of the WT1 & 2 parents and any F2 recombinants, and identify putative clones with the expected mobility.
9) Pick colonies for 4-6 putative clones from the plate generated in step 2 above and generate O/N cultures for plasmid isolation and Southern blot analysis (see below).
Clone verification II: Southern
1) Isolate plasmid DNA (by any standard technique) from O/N cultures of clones selected in step 9 above.
2) PCR label plasmid DNA by linear amplification using the MseI primer (used in step 1 of Sec. 4.1 ) as described below.
Single strand PCR labeling (recipe is for a single labeling rxn)- amplify using "PRE-AFLP" theromcycle profile
3 µL MseI-primer (@ 50 ng/µL)
1.0 µL 0.5 mM dATP, dGTP, dTTP
15.0 µL a32P-dCTP (3000 Ci/mmol)
2.5 µL 10x-Promega Taq buffer
1.5 µL MgCl2
0.5 µL Taq Pol
1.0 µL dH2O
1.0 µL ds plasmid DNA (25 ng/µL)
3) ElectroBlot a standard AFLP gel (according to standard methods eg. Ausubel et al Current Protocols in Molecular Biology)of WT1 & WT2 parents, any recombinants having the WT2-specific band, and a few non-recombinants F2s (ones not having the WT2-specific band; as controls), that have been amplified as per Sec. 2.0-2.2 using the EcoRI- (33P end-labeled) and MseI-primers that were used in step 1 of Sec. 4.1.4.3 Fragments and oligos for library screening4) Hybridize w/ labeled plasmid by standard methods.
[WT2-specific clones can be unambiguously determined from an autorad of the hybridized blot.]
- Cloned AFLP fragments can be used in a variety of manners as tools to screen various nucleic acid libraries (YAC, BAC, P1, cosmid, lambda, cDNA), a couple of examples are given below: -AcknowledgmentsSequence based probes
1) Sequence cloned AFLP fragment(s) by standard methods.
2) Based on sequence, generate oligos to sites internal to the EcoRI and MseI adapter sequences (plant specific sequences).
3) Use these oligos to screen various libraries via PCR.
Insert based probes
- Use the insert or whole plasmid containing the of interest in a variety of ways -
1) Label ds insert DNA that has been liberated from vector DNA by standard RE digestion. Use labeled DNA in hybridizations for filter based library screening by standard techniques.
2) Label whole, linearized ds plasmid containing the insert and use it for screening filters as above in step 1.
3) Use oligos generated in step 2 of Sequence based probes above to make single stranded insert DNA probes. This can be accomplished as described in step 2 of the Clone verification II section of Sec. 4.2, with the exception that both insert-specific oligos are used as primers instead of an MseI-primer (adjust total vol appropriately). Use this labeled DNA for screening filter as described in step 1 above.
We wish to thank Dr. Chris Somerville for introducing us to the existence of AFLP technology and for his continued support and help throughout our work. We also wish to thank Dr. Shuana Somerville and members of her laboratory for many helpful discussions. Finally, we wish to thank Dr. Winslow Briggs who has embraced this technology and provided financial support for much of the AFLP work ongoing in the lab. We are also indebted to Glenn Ford for his assistance in providing this information in appropriate WWW formats.
Literature Cited
Liscum E and Briggs WR (1995) Mutations in the NPH1 locus of Arabidopsis disrupt the perception of phototropic stimuli. Plant Cell 7: 473-485.
Thomas CM, Vos P, Zabeau M, Jones DA, Norcott KA, Chadwick BP and Jones JDG (1995) Identification of amplified restriction fragment polymorphism (AFLP) markers tightly linked to the tomato Cf-9 gene for resistance to Claosporium fulvum. Plant J 8: 785-794.
Vos P, Hogers R, Bleeker M, Reijans M, van de Lee T, Hornes M, Frijters A, Pot J, Peleman J, Kuiper M and Zabeau M (1995) AFLP: a new technique for DNA fingerprinting. Nucl. Acids Res. 23: 4407-4414.
Zabeau M (1992) European Patent Application, publication no. EP 0534858
Zabeau M and Vos P (1993) European Patent Application, publication no. EP 0534858.