【精华】vol 687 Chapter 4 巢式PCR侧翼序列测定实验
丁香园

下面的是一个巢式PCR测定实例,更多分享,找学霸师兄哦,师兄微信号:shixiongcoming
Methods in Molecular Biology vol.687 Park D.J. (ed.) PCR Protocols Chapter 4
PART II :Cloning and Sequencing
Lariat-Dependent Nested PCR for Flanking Sequence Determination
Daniel J. Park
Abstract
Methods detailed in this chapter relate to the use of Lariat-dependent Nested (LaNe) PCR to characterize unknown RNA or DNA sequence flanking known regions. A multitude of approaches designed to determine flanking sequences have been described in the literature. Variously, problemsrelated to these approaches include lack of resolution or failure, depending on experimental context, and complex handling. LaNe-based methods are designed to harness “two-sided” genespecific PCR with the option of nesting but without the requirement for inefficient and involvedenzyme preprocessing steps.
Key words: Flanking sequence determination, LaNe, PCR walking
1. Introduction
While relatively recent higher throughput DNA sequencing isrevolutionizing the way biological research is conducted, thereremain applications for more focused analyses of flankingsequence. The size of the region to be resolved, number of specimens,complexity and duration of processing and bioinformaticanalysis, intractability of sequence to methodology, and cost areamong the factors influencing the choice of approach to the studyof flanking sequence. Many methods for PCR-based flankingsequence determination have been described, such as 3?RapidAmplification of cDNA Ends (3?RACE), 5?RACE, RapidAmplification of Genomic DNA Ends, Inverse PCR, CapSelect,Ligase-mediated PCR, Splinkerette PCR, Thermal AsymmetricInterlaced (TAIL) PCR, and Universal Fast Walking (1–11). Tonooka and Fujishima have recently reviewed PCR methods for walking along genomic DNA (12). Methods for PCR-basedflanking sequence determination exhibit various limitations,including a requirement for relatively large quantities of inputmaterial, complex processing steps relying on multiple handlingsteps, and dependence on efficient enzyme steps involving restrictionendonucleases, ligases, and/or exonuclease processing. Inmost cases, there is a reliance on the so-called “one-sided” PCRin which extension is primed by a gene-specific primer and a“general” primer targeted to sites common across the genome oradapters present in most species of the template mix. Theseapproaches work satisfactorily in many cases, but some canencounter difficulties in more challenging contexts. In particular,one-sided approaches often yield “smeared products” as visualizedfollowing gel electrophoresis such that desired products ofinterest are difficult to resolve.
The Lariat-dependent Nested (LaNe)-based methods 3?RACELaNe, 5?RACE LaNe, and LaNe RAGE (13–15) are derived fromthe “panhandle” concept employed in Universal Fast Walking(UFW) in which a lariat structure is involved in self-priming toyield a template which enables “two-sided” and nested PCR to beapplied to flanking sequence determination. LaNe-based methodsstreamline the UFW concept workflow and system by removingthe requirement for multiple rounds of exonuclease addition and“end-fill” steps prior to initiating “PCR-proper.” Instead, LaNebasedself-priming is facilitated by denaturation and annealingsteps inherent to conventional PCR thermocycling (the reactionscheme for LaNe RAGE is depicted in Fig. 1).
In this chapter, LaNe-based protocols are presented for 5? or3? oriented RNA flanking sequence determination (5?RACELaNe and 3?RACE LaNe) and genomic DNA flanking sequencedetermination (LaNe RAGE). Further, special requirements andtips for customization of the approaches are presented inSubheading 4.
2. Materials
2.1. 3' RACE LaNe
1. 20 ng total RNA (see Note 1).
2. 10 mM dNTPs.
3. ThermoScript ® Reverse Transcriptase and accompanyingbuffers (Invitrogen) (see Note 2).
4. TTT3 primer: 5'-gene-specific sequence followed byTTTTTTTTTTTTTTTTV-3?. The 5-prime gene-specificregion of this primer should be in the same orientation asGSP1 and GSP4 (see Subheading 2.1, item 7) in alignmentwith position “3” as depicted in Fig. 1.
5. RNaseH (Invitrogen).

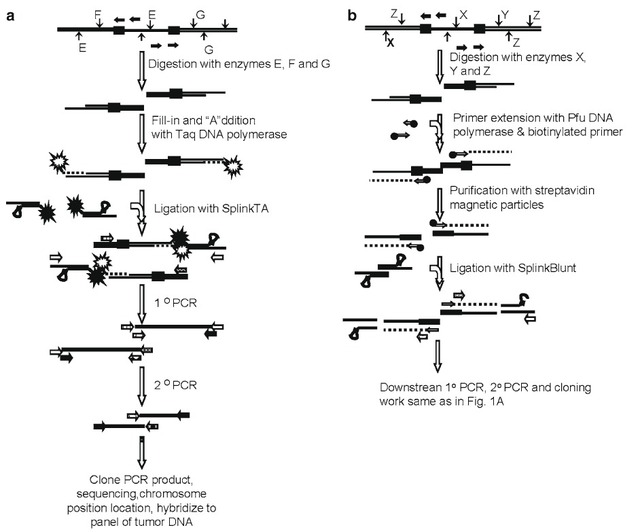
Fig. 1. Schematic illustration of the LaNe approach applied in LaNe RAGE. A hybridprimer containing gene-specific sequence at the 5? end and nongene-specific sequenceat the 3? end is used to prime extension from the template (hybrid primer extension). Theresulting product is then amenable to gene-specific second strand synthesis, followedby lariat structure formation and self-primed extension, to yield template which can beamplified by “two-sided PCR” using two gene-specific primers (Single tube lariatdependentPCR). This can occur in a standard PCR thermocycling profile due to themultiple rounds of denaturation and annealing in the presence of the appropriate primersand thermostable DNA polymerase. The product of Single-tube lariat-dependent PCRcan be applied as template in a nested PCR scheme (optional nested PCR). Numbersindicate features of a known sequence region. It is important to note that features 1–6should always be oriented with feature 6 closest to the unknown sequence to be determined. The dashed line indicates the unknown sequence of the template which is to bedetermined. Underlined numbers indicate reverse complement sequences with respectto these features. Primers are represented by horizontal arrows. Dotted lines indicatepolymerase extension.
6. PfuTurbo ® DNA Polymerase and accompanying buffers andsolutions (Stratagene) (see Note 3).
7. GSP1 and GSP4: Gene-specific primers 1 and 4 should bedesigned according to standard PCR primer conventions withrespect to avoiding primer dimers and intraprimer annealing,attempting to prevent nonspecific priming, and ensuring thatthe Tm of the primers is a few degrees higher than the intendedannealing temperature during thermal cycling but that both arein the same orientation as each other with the 3? ends pointingtowards the unknown region to be elucidated (refer to Fig. 1)(see Notes 4 and 5). GSP1 and GSP4 should be designed inalignment with positions “1” and “4,” respectively, in Fig. 1.
8. GSP2 and GSP5: Gene-specific primers (optional – see Notes 6and 7) should be designed in a similar manner to GSP1 and GSP4but aligned with positions “2” and “5,” respectively, in Fig. 1.
9. QIAEXII DNA purification system (Qiagen) (optional –see Note 8).
10. GSP6: Gene-specific primer (optional – see Note 8) shouldbe designed in a similar manner to GSP1, GSP2, GSP4, andGSP5 but aligned with position “6” in Fig. 1.
2.2. 5 ' RACE LaNe
2.2.1. First-Strand Intrastrand-Annealing 5' RACE LaNe (FI RACE LaNe)
1. 200 ng total RNA (see Note 1).
2. 10 mM dNTPs.
3. ThermoScript? Reverse Transcriptase and accompanyingbuffers (Invitrogen) (see Note 2).
4. RT primer: Reverse transcription primer should be orientedsuch that the 3-prime end is directed toward the unknown5-prime mRNA region to be elucidated and reverse transcribedproduct includes features “1” to “6” of Fig. 1.
5. RNaseH (Invitrogen).
6. PfuTurbo®DNA Polymerase and accompanying buffers andsolutions (Stratagene) (see Note 3).
7. GSP1 and GSP4: Gene-specific primers 1 and 4 (seeSubheading 2.1, item 7 and Fig. 1). In this case, 3-prime endsof primers are all oriented towards the unknown 5-prime regionof mRNA to be elucidated.
8. GSP2 and GSP5: Gene-specific primers 2 and 5 (seeSubheading 2.1, item 8 and Fig. 1). In this case, 3-prime endsof primers are all oriented towards the unknown 5-primeregion of mRNA to be elucidated.
9. QIAEXII DNA purification system (Qiagen) (optional – seeNote 8).
10. GSP6: Gene-specific primer 6 (see Subheading 2.1, item 10and Fig. 1) (optional – see Note 8). In this case, 3-prime endsof primers are all oriented towards the unknown 5-primeregion of mRNA to be elucidated.
2.2.2. Termina Transferase-Mediate 5‘ RACE LaNe (TT RACE LaNe)
1. 200 ng total RNA (see Note 1).
2. 10 mM dNTPs.
3. SuperScript? II Reverse Transcriptase and accompanyingbuffers (Invitrogen) (see Note 9).
4. 25 mM manganese chloride.
5. 5 mg/ml bovine serum albumin.
6. RT primer: Reverse transcription primer should be orientedsuch that the 3-prime end is directed toward the unknown5-prime mRNA region to be elucidated and reverse transcribedproduct includes features “1” to “6” of Fig. 1.
7. RNaseH (Invitrogen).
8. QIAEXII DNA purification system (Qiagen).
1. 200 ng total RNA (see Note 1).
2. 10 mM dNTPs.
3. SuperScript? II Reverse Transcriptase and accompanyingbuffers (Invitrogen) (see Note 9).
4. 25 mM manganese chloride.
5. 5 mg/ml bovine serum albumin.
6. RT primer: Reverse transcription primer should be orientedsuch that the 3-prime end is directed toward the unknown5-prime mRNA region to be elucidated and reverse transcribedproduct includes features “1” to “6” of Fig. 1.
7. RNaseH (Invitrogen).
8. QIAEXII DNA purification system (Qiagen).
2.3. LaNe RAGE
1. 300 ng genomic DNA (see Note 1).
2. 10 mM dNTPs.
3. PfuTurbo?DNA Polymerase and accompanying buffers andsolutions (Stratagene) (see Note 3).
4. NNN3: First-strand synthesis primer 5?-gene-specific sequencefollowed by NNNNNCTCAC-3? (see Note 10). The 5-primegene-specific region of this primer should be in the same orientationas GSP1 and GSP4 (see Subheading 2.3, item 5) inalignment with position “3” as depicted in Fig. 1.
5. GSP1 and GSP4: Gene-specific primers 1 and 4 (seeSubheading 2.1, item 7 and Fig. 1). In this case, 3-primeends of primers are all oriented towards the unknown regionof genomic DNA to be elucidated.
6. GSP2 and GSP5: gene-specific primers 2 and 5 (seeSubheading 2.1, item 8 and Fig. 1). In this case, 3-primeends of primers are all oriented towards the unknown regionof genomic DNA to be elucidated.
7. QIAEXII DNA purification system (Qiagen) (optional – seeNote 8).
8. GSP6: Gene-specific primer 6 (see Subheading 2.1, item 10and Fig. 1) (optional – see Note 8). In this case, 3-prime endsof primers are all oriented towards the unknown region ofgenomic DNA to be elucidated.
3. Methods
LaNe-based methods represent a suite of tools that are applicableto the elucidation of 5-prime and 3-prime mRNA flankingsequence (5?RACE LaNe and 3?RACE LaNe, respectively) andgenomic DNA flanking sequence in either direction (LaNeRAGE). Theoretically, the latter could be applied to the determinationof insertion elements such as transposons as part of mutagenesisscreening or DNA fingerprinting strategies, althoughthese applications remain to be tested. The examples that followrepresent demonstration methods that have been tested. It shouldbe noted that elements of the respective approaches could beapplicable to alternative approaches within the suite of tools (seeNotes 11 and 12).
3.1. 3' RACE LaNe
The reaction scheme for 3?RACE LaNe is similar to that depictedin Fig. 1 for LaNe RAGE, except NNN3 is swapped for thereverse transcription primer TTT3 in the first step.
1. TTT3 primer (1 mM final concentration) is used to primeThermoScript?-catalysed reverse transcription of 20 ng totalRNA at 55°C for 1 h in a total reaction volume of 20 ml.
2. Reverse transcription products are treated with 2 U RNaseH at37°C for 20 min prior to heat inactivation at 85°C for 10 min.
3. Five microliters of this product are used to template a 50 mlPfuTurbo?hot start PCR in (see Note 13) 1× reaction bufferincluding 300 mM dNTPs, 3.75 U PfuTurbo?DNA
Polymerase, and 250 nM GSP1 and GSP4. Thermocyclingparameters used are: 40 cycles of 94°C for 1 min, 50°C for1 min, 72°C for 5 min (see Note 7).
4. Product can be analyzed by agarose gel electrophoresis at thisstage (see Note 6). Single band or multiple banding profilesmight be expected depending on experimental conditions(see Note 14). Optionally, a nested PCR can be performed,in which case, proceed to the next step.
5. Use 0.5 ml of product from the preceding step to template anested 50 ml PfuTurbo?hot start PCR in (see Note 13) 1×reaction buffer including 300 mM dNTPs, 3.75 U PfuTurbo® DNA Polymerase, and 250 nM GSP2 and GSP5. Thermocycling parameters used are: 40 cycles of 94°C for1 min, 58°C for 1 min, 72°C for 5 min (see Note 7).
6. Product can be analyzed by agarose gel electrophoresis at thisstage. Single band or multiple banding profiles might be expecteddepending on experimental conditions (see Note 14).
7. Product can be subjected to direct Sanger sequencing usingGSP5 or GSP6 as the sequencing primer (see Note 8).
8. Selected post-agarose gel electrophoresis resolved productscan be gel-extracted using the QIAEXII gel extractionsystem, followed by direct Sanger sequencing using GSP5 orGSP6 as the sequencing primer (see Note 8).
3.2. 5' RACE LaNe
Two example protocols are presented: FI RACE LaNe and TTRACE LaNe. The former uses a relatively streamlined workflowbut a less “defined” mechanism for lariat structure formation (seeNote 2). As such, FI RACE LaNe may be useful in some contextsbut not others. The first step of FI RACE LaNe employs a nonhybridreverse transcription primer instead of NNN3 depicted inFig. 1 and relies on fortuitous lariat structure formation andextension by the first DNA strand. Ensure that features 1–6 withreference to Fig. 1 are oriented such that feature 6 is closest to theunknown 5-prime sequence. TT RACE LaNe is designed to controllariat structure formation to facilitate a more robust approachbut uses a more cumbersome protocol to achieve this. Similarly,ensure that features 1 to 6 are oriented such that feature 6 is closestto the unknown 5-prime sequence. Reverse transcription andterminal transferase processing precede priming by GSPT12G3,which is analogous to NNN3 priming depicted in Fig. 1.
3.2.1. FI RACE LaNe
1. RT primer (1 mM final concentration) is used to primeThermoScript?-catalyzed reverse transcription of 200 ng totalRNA at 50°C for 1 h in a total reaction volume of 20 ml.
2. Reverse transcription products are treated with 2 U RNaseH at37°C for 20 min prior to heat inactivation at 85°C for 10 min.
3. Two microliters of this product are used to template a 50 mlPfuTurbo?hot start PCR in (see Note 13) 1× reaction bufferincluding 300 mM dNTPs, 3.75 U PfuTurbo?DNA
Polymerase, and 250 nM GSP1 and GSP4. Thermocyclingparameters used are: 40 cycles of 94°C for 1 min, 55°C for1 min, 72°C for 2 min (see Note 7).
4. Product can be analyzed by agarose gel electrophoresis at thisstage (see Note 6). Single band or multiple banding profilesmight be expected depending on experimental conditions(see Note 14). Optionally, a nested PCR can be performed,in which case, proceed to the next step.
5. Use 0.5 ml of product from the preceding step to template anested 50 ml PfuTurbo?hot start PCR in (see Note 13)1× reaction buffer including 300 mM dNTPs, 3.75 UPfuTurbo?DNA Polymerase, and 250 nM GSP2 and GSP5. Thermocycling parameters used are: 40 cycles of 94°C for1 min, 60°C for 1 min, 72°C for 2 min (see Note 7).
6. Product can analyzed by agarose gel electrophoresis at this stage. Single band or multiple banding profiles might be expecteddepending on experimental conditions (see Note 14).
7. Product can be subjected to direct Sanger sequencing usingGSP5 or GSP6 as the sequencing primer (see Note 8).
8. Selected post-agarose gel electrophoresis resolved productscan be gel-extracted using the QIAEXII gel extraction system,followed by direct Sanger sequencing using GSP5 orGSP6 as the sequencing primer (see Note 8).
3.2.2. TT RACE LaNe
1. RT primer (1 mM final concentration) is used to primeSuperScript? II-catalyzed reverse transcription of 200 ngtotal RNA in 1× buffer in the presence of 2 mM manganesechloride and 0.1 mg/ml bovine serum albumin at 42°C for1 h in a total reaction volume of 20 ml (see Note 9).
2. Reverse transcription products are treated with 2 U RNaseH at37°C for 20 min prior to heat inactivation at 85°C for 10 min.
3. Products are cleaned using the QIAEXII DNA purificationsystem and eluted using 20 ml of elution buffer. 15 ml of eluateis collected, with care taken to avoid transfer of matrixslurry to the collection tube.
4. Five microliters of eluate is included in a 20 ml terminal transferasereaction in 1× buffer with the inclusion of 200 uMdATP and 20 U terminal transferase at 37°C for 20 min.
5. Two microliters of this product are used to template a 50 ulPfuTurbo?hot start PCR in (see Note 13) 1× reaction bufferincluding 300 uM dNTPs, 3.75 U PfuTurbo® DNA
Polymerase, 100 nM GSPT12G3, and 250 nM GSP1 andGSP4. Thermocycling parameters used are: 40 cycles of 94°Cfor 1 min, 55°C for 1 min, 72°C for 2 min (see Note 7).
6. Product can be analyzed by agarose gel electrophoresis at thisstage (see Note 6). Single band or multiple banding profilesmight be expected depending on experimental conditions(see Note 14). Optionally, a nested PCR can be performed,in which case, proceed to the next step.
7. Use 0.5 ul of product from the preceding step to template anested 50 ul PfuTurbo®hot start PCR in (see Note 13) 1×reaction buffer including 300 uM dNTPs, 3.75 U PfuTurbo® DNA Polymerase, and 250 nM GSP2 and GSP5. Thermocycling parameters used are: 40 cycles of 94°C for1 min, 60°C for 1 min, 72°C for 2 min (see Note 7).
8. Product can analyzed by agarose gel electrophoresis at this stage.
Single band or multiple banding profiles might be expecteddepending on experimental conditions (see Note 14).
9. Product can be subjected to direct Sanger sequencing usingGSP5 or GSP6 as the sequencing primer (see Note 8).
10. Selected post-agarose gel electrophoresis resolved productscan be gel-extracted using the QIAEXII gel extraction system, followed by direct Sanger sequencing using GSP5 orGSP6 as the sequencing primer (see Note 8).
3.3. LaNe RAGE
Figure 1 schematically depicts the steps in LaNe RAGE. It mustbe ensured that the orientation of features 1–6 are such that feature6 is closest to the unknown region to be characterized.
1. 50 ul 1× PfuTurbo?buffer containing 300 ng genomic DNAand 250 nM NNN3 is heated to 94°C for 3 min and thenplaced on ice for 5 min prior to the inclusion of 3.75 UPfuTurbo® and dNTPs to achieve a concentration of 300 mM.
2. Heat the reaction tubes from 18°C to 72°C at a rate of 0.1°Cper 5 s and hold at 72°C for 10 min prior to placing on ice(see Note 10).
3. Include GSP1 and GSP4 to achieve 250 nM concentrationsof each and proceed immediately to thermocycling.
Thermocycling parameters used are: 40 cycles of 94°C for1 min, 58°C for 1 min, 72°C for 2 min (see Note 7).
4. Product can be analysed by agarose gel electrophoresis at thisstage (see Note 6). Single band or multiple banding profilesmight be expected depending on experimental conditions(see Note 7). Optionally, a nested PCR can be performed, inwhich case, proceed to the next step.
5. Use 0.5 ul of product from the preceding step to template anested 50 ml PfuTurbo?hot start PCR in (see Note 13) 1×reaction buffer including 300 uM dNTPs, 3.75 U PfuTurbo® DNA Polymerase, and 250 nM GSP2 and GSP5. Thermocycling parameters used are: 40 cycles of 94°C for1 min, 60°C for 1 min, 72°C for 2 min (see Note 7).
6. Product can analysed by agarose gel electrophoresis at this stage.Single band or multiple banding profiles might be expecteddepending on experimental conditions (see Note 14).
7. Product can be subjected to direct Sanger sequencing usingGSP5 or GSP6 as the sequencing primer (see Note 8).
8. Selected post-agarose gel electrophoresis resolved productscan be gel-extracted using the QIAEXII gel extraction system,followed by direct Sanger sequencing using GSP5 orGSP6 as the sequencing primer (see Note 8).
4. Notes
1. The integrity of the input nucleic acid specimen will have a considerablebearing on the potential for success using LaNe-basedapproaches, as with most molecular biology technologies. If theinput material is significantly degraded or fragmented, only
relatively short regions of flanking sequence may be resolvable inany given “LaNe step.” In this context, it is perhaps unwise to designprimers and conditions in an attempt to yield long products.
2. ThermoScript? Reverse Transcriptase is relatively thermostable,permitting relatively stringent priming of reverse transcription,if required. Alternative enzymes with similarproperties should also be suitable.
3. Terminal transferase activity is undesirable in a thermostableDNA-dependent DNA polymerase employed in LaNe-basedmethods in which the lariat structure formation is facilitatedby a gene-specific hybrid primer sequence incorporationevent, e.g., via TTT3, GSPT12G3, or NNN3. Such activitywould result in a majority of would-be lariat-primed eventsexhibiting mismatches at the 3-prime end, thus inhibitinglariat-primed extension dramatically. This does not apply toexamples such as FI RACE LaNe or in other cases in whichthe principle of “fortuitous” lariat structure priming isapplied (e.g., in principle, an iteration of LaNe RAGE couldremove the requirement for NNN3-primed strand synthesisto simply employ nested PCR using GSP1 and GSP4, followedby GSP2 and GSP5 – see Notes 6 and 12). PfuTurbo? DNA Polymerase has been used effectively, but alternativeenzymes with similar properties should also suit LaNe-basedmethods.
4. In the LaNe-based methods, for all primers used, gene-specificprimer regions are always oriented with the 3-prime endtowards the unknown sequence to be elucidated. In caseswhere hybrid primer is used, e.g., TTT3, GSPT12G3, andNNN3, the gene-specific region is always located at the5-prime end of the primer, but the orientation of the genespecificprimer sequence is designed with its 3-prime end orientedtowards the unknown sequence to be elucidated as withall other primers. This works because end-filling and subsequentlariat-based priming (as part of the LaNe mechanism)yield reverse complement forms of some sites at one end ofthe template, allowing primers to engage in two-sided PCR.
5. Primer design software such as OligoAnalyzer 3.1, Primer 3,and Primer-BLAST can be employed to design primersaccording to the desired melting temperature (aim for “typical” PCR primer properties) and to check for and avoid primersor primer combinations liable to form strong secondarystructures (e.g., hairpins) or engage in primer dimer formation. PCR primers can be filtered against repetitive elementsof reference genomes and checked for predicted off-targetproducts. The scope for primer design is somewhat dependent,however, on the amount of known sequence availableand the desire to design the primers to anneal close to theunknown region to be resolved if possible.
6. In some applications, just one round of PCR may be sufficient.
This requires fewer primers and a shorter region of knownsequence to design LaNe primers. There will be other caseswhere a single round of PCR is insufficient, and the addedsensitivity and specificity afforded by a second nested PCRwill resolve desired products.
7. The RT and PCR conditions detailed in this chapter representproof-of-concept methodologies. These are most likelyopen to considerable optimization to suit requirements. Forexample, some applications may require greater or lowerstringency necessitating higher or lower annealing temperatures,respectively. Some applications may require longerextension times during cycling to facilitate yield of longerproducts. Applications may require more or less enzyme orinitial nucleic acid template. It is likely that fewer PCR cycleswould be necessary than are detailed in this chapter. However,it should be noted that by their nature, LaNe-based protocolsemploy templates that form suppressive “panhandle” structures. As such, relatively high concentrations of primers GSP1and GSP4 or GSP2 and GSP5 may be required to competeeffectively with such structure formation for effective primingduring thermocycling. Ideally, the primer regions should berelatively close together to minimize the relative stability andrate of formation of such structure formation (primer sitescould even overlap to a limited extent). Conceivably, GSP1and GSP4 or GSP2 and GSP5 could employ “heel clamps” toincrease Tm and improve competitivity.
8. As a fast and convenient approach, products can be sequenceddirectly using GSP5 or GSP6 as the sequencing primer. Insome cases, it may be desirable or necessary to isolate productbands from an agarose gel. Following gel extraction, productscan be separately sequenced using GSP5 or GSP6 as thesequencing primer. Ideally, sequencing primers should be setback sufficiently from the unknown flanking sequence understudy to allow verification of gene-specific amplicon. If theformer is not possible due to design constraints or if preferred,product can be cloned into a plasmid and sequencing performedusing plasmid-specific sequencing primers.
9. TT RACE LaNe makes use of the inherent terminal transferaseactivity of M-MLV-based reverse transcriptase, e.g.,SuperScript? II or Expand-RT, in the presence of manganesechloride as an additional divalent cation to magnesium chlorideand in the presence of BSA as a stabilizing agent. Underthese conditions, several cytosine residues are typically addedin the presence of 5? capped mRNA templates. In the presenceof 5?-OH or 5?-phosphate termini, the activity is lesspronounced and not cytosine-specific (3).
10. As a proof-of-concept utility, NNN3 was designed with a“NNNNNCTCAC” 3-prime terminus. However, any numberof variations could be used to bias towards more frequentor less frequent hybridization sites to influence the tendencyof product sizes. For instance, a longer “defined” region atthe 3-prime end might tend towards the yield of longer products,since hybrid primer binding sites would be less frequent. The nature of the defined sequence at the 3-prime end couldbe varied to suit the experimental context. There is considerablescope to influence the performance of the system byadjusting hybrid primer sequence. Integrally related to this isthe way in which annealing of NNN3 to denatured genomicDNA is conducted prior to extension. More stringent conditionsshould result in fewer hybrid primer binding events andfewer products, to enable the yield of longer products. Forexample, immediately following denaturation (in the presenceof template, NNN3, PfuTurbo?DNA Polymerase, anddNTPs in reaction buffer) one might perform annealing at anelevated temperature, e.g., 45–55°C, for several minutes(without having cooled below this temperature) prior toslowly raising the temperature to 72°C (e.g., 1°C/20 s tobalance proficiency of extension at suboptimal temperatureswith maintenance of annealing), with all subsequent primersadded at this temperature. Further, a reduction in concentrationof NNN3 may contribute to less frequent hybrid primerbinding events to permit “cleaner” product profiles and selectionof longer products.
11. It is likely that LaNe RAGE would be useful in applicationssuch as transposon insertional mutagenesis screening programsto identify insertion sites of mutants and in DNA fingerprinting,as has been demonstrated using Universal Fast Walking. However, these experiments have not been conducted.
12. One approach that may be valuable but has not been testedto date would combine aspects of CapTrapper (16) and5?RACE LaNe, in which 5-prime capped mRNAs would bephysically separated from noncapped mRNA species prior toutilization of a LaNe-based approach. Fortuitous lariat structureformation might be employed in LaNe RAGE withoutfirst-strand synthesis primed by NNN3, analogous to thatused in TT RACE LaNe. This would result in a less-controlledbut more streamlined protocol in which the user couldproceed directly to a hot-start PCR templated with genomicDNA using primers GSP1 and GSP4, optionally followed bya nested PCR using GSP2 and GSP5.
13. In the proof-of-concept methods detailed, hot-start PCR wasperformed by separate addition of enzyme following thermal denaturation. Alternatively (and preferably), chemical orantibody inhibition-based hot-start enzyme may be employed.
14. Product profiles vary considerably and are strongly influencedby hybridization conditions, primer design, and PCR parametersas discussed in this section.
Acknowledgments
This work was supported by the National Health and MedicalResearch Council of Australia grant no. 148630 to ProfessorsJ.A.M. Graves and M.B. Renfree, Associate Professor V.R. Harleyand Dr. A.J. Pask.
References
1. Frohman, M.A., Dush, M.K., and Martin, G.R. (1988) Rapid production of full-length cDNAs from rare transcripts: amplification using a singlegene-specific oligonucleotide primer. Proc. Natl. Acad. Sci. USA 85, 8998–9002.
2. Frohman, M.A. and Martin, G.R. (1989) Rapid amplification of cDNA ends using nested primers. Technique 1, 165–70.
3. Schmidt, W.M. and Mueller, M.W. (1999) CapSelect: a highly sensitive method for 5'CAP-dependent enrichment of full-length cDNA in PCR-mediated analysis of mRNAs.Nucleic Acids Res. 27, e31.
4. Maruyama, K. and Sugano, S. (1994) Oligocapping: a simple method to replace the cap structure of eukaryotic mRNAs with oligoribonucleotides. Gene 138, 171–4.
5. Volloch, V., Schweitzer, B., and Rits, S. (1994)Ligation-mediated amplification of RNA from murine erythroid cells reveals a novel class ofbeta globin mRNA with an extended 5'-untranslated region. Nucleic Acids Res. 2,2507–11.
6. Cormack, R.S. and Somssich, I.E. (1997) Rapid amplification of genomic ends (RAGE) as a simple method to clone flanking genomic DNA. Gene 194, 273–6.
7. Ochman, H., Gerber, A.S., and Hartl, D.L. (1988) Genetic applications of an inverse polymerase chain reaction. Genetics 120, 621–3.
8. Mueller, P.R. and Wold, B. (1989) In vivo footprinting of a muscle specific enhancer byligation mediated PCR. Science 246, 780–6.
9. Devon, R.S., Porteous, D.J., and Brookes, A.J.(1995) Splinkerettes – improved vectorettes for greater efficiency in PCR walking. NucleicAcids Res. 23, 1644–5.
10. Liu, Y.-G. and Whittier, R.F. (1995) Thermalasymmetric interlaced PCR: automatable amplification and sequencing of insert end fragments from P1 and YAC clones for chromosomewalking. Genomics 25, 674–81.
11. Myrick, K.V. and Gelbart, W.M. (2002) Universal fast walking for direct and versatiledetermination of flanking sequence. Gene 284, 125–31.
12. Tonooka, Y. and Fujishima, M. (2009) Comparison and critical evaluation of PCRmediatedmethods to walk along the sequence of genomic DNA. Appl. Microbiol. Biotechnol. 85, 37–43.
13. Park, D.J. (2004) 3?RACE LaNe: a simple andrapid fully nested PCR method to determine 3' -terminal cDNA sequence. Biotechniques 36, 586–90.
14. Park, D.J. (2005) A new 5? terminal murineGAPDH exon identified using 5?RACE LaNe. Mol. Biotechnol. 29, 39–45.
15. Park, D.J. (2005) LaNe RAGE: a new tool forgenomic DNA flanking sequence determination. Electronic J. Biotechnol. 8, 218–25.
16. Carninci, P., Westover, A., Nishiyama, Y.,Ohsumi, T., Itoh, M., Nagaoka, S., Sasaki, N.,Okazaki, Y., Murumatsu, M., Schneider, C., andHayashizaki, Y. (1997) High efficiency selectionof full-length cDNA by improved biotinylated cap trapper. DNA Res. 4, 61–6.