




师从施一公,发表 6 篇Science/3篇 Cell,她被誉为「世界最具潜力女科学家」| 论文盘点
丁香学术
10 月 31 日,2022 达摩院青橙奖公布,15 位平均年龄仅 33 岁的中国青年科学家获奖,每位获奖者将得到百万奖金及科研支持。获奖者中有 2 位从事生命科学领域,其中一位是来自西湖大学生命科学学院的副研究员白蕊。
她还曾入选由中国科协评选的「未来女科学家计划」,获得欧莱雅-联合国教科文组织评选的「世界最具潜力女科学家奖」。

图片来源:西湖大学官网
白蕊本科就读于武汉大学,在大三时听了施一公的一场讲座后,被他的科研观和大局观吸引,坚持选施一公作为导师。此后,便接触到了「剪接体」这一世界级难题。
RNA 剪接是生命体解读遗传密码的核心步骤,即把遗传密码中的内含子「剪」出来,外显子「接」一起的过程,由细胞核内的剪接体负责执行。人类的遗传疾病大约有 35% 都是因为剪接异常造成的。然而,剪接体催化过程中不同构象高分辨率结构的缺失严重限制了大家对其工作机制以及 RNA 剪接的分子机理的理解。因此,对于剪接体以及 RNA 剪接通路上各复合物结构的研究,是当今世界最富有挑战性、最亟待解决的课题之一。
她曾以第一作者和共同第一作者的身份在 Science 上发表 6 篇论文,在 Cell 上发表论文 3 篇。现在的白蕊参与并主导了全球唯一覆盖完整 RNA 循环的剪接体系列成果,为相关遗传病和癌症机理研究带来新思路。
白蕊的研究成果,也以封面的形式写进了国际最权威的生化教科书里。
施一公实验室报道的首个酵母剪接体的结构
图片来源:生物化学经典教材 Lehninger Principles of Biochemistry(第七版)封面
今天,我们来简单盘点一下近年来白蕊的 10 篇科研成果。(以时间顺序排列)
1. The 3.8 Å structure of the U4 /U6.U5 tri-snRNP: Insights into spliceosome assembly and catalysis
(Science , 29 Jan 2016)
(图片来源:Science)
前体信使 RNA 的剪接是由称为剪接体的动态巨型复合体完成的。功能性剪接体的组装需要一个预先组装的 U4 /U6.U5tri-snRNP 复合物,它由 U5 小核核糖核蛋白(SnRNP)、U4 和 U6 小核 RNA(SnRNA)双链以及一系列蛋白质因子组成。
该研究报道了一株酿酒酵母 U4 /U6.U5tri-snRNP 的三维结构,其总分辨率为 3.8 埃。Tri-snRNP 核心区的局部分辨率达到 3.0 到 3.5 埃,可以构建精细的原子模型。
作者解析的结构包括 U5snRNA、广泛碱基配对的 U4 /U6snRNA 以及包括 Prp8 和 Snu114 在内的 30 个蛋白质,共计 8495 个氨基酸和 263 个核苷酸,总分子质量约为 100 万道尔顿。
来自 U6snRNA 的催化核苷酸 U80 以非活性构象存在,由其与 U4snRNA 的碱基对稳定相互作用,并受 Prp3 保护。前信使 RNA 通过与 U6snRNA 和 U5snRNA 的环 I 的碱基配对相互作用结合在 Tri-snRNP 中。这种结构与剪接体的结构一起揭示了剪接体核酶激活过程中 snRNAs 的分子编排。
酿酒酵母 U4 /U6.U5tri-snRNP 剪接体的结构(图片来源:Science)
2. Structure of a yeast activated spliceosome at 3.5 Å resolution
(Science 26 Aug 2016)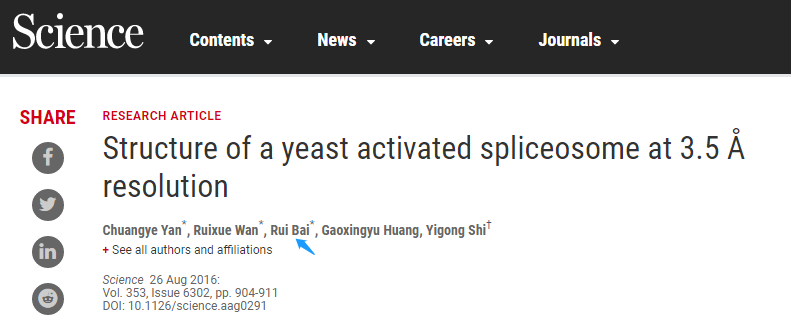
(图片来源:Science)
这篇研究报道了来自酿酒酵母的一个激活态剪接体(称为 Bact 复合体)的原子结构,它是由平均分辨率为 3.52 埃的冷冻电子显微镜测定的。
最终的精制模型包含 U2 和 U5 小核核糖核蛋白颗粒(SnRNPs),U6 小核 RNA(SnRNA),19 个复合物(NTC),NTC 相关(NTR)蛋白,以及一个 71 核苷酸的 Pre-mRNA 分子,共计来自 38 种蛋白质的 13505 个氨基酸,总分子量约为 160 万道尔顿。
5ʹ外显子由 U5snRNA 的环 I 锚定,而内含子的 5ʹ剪接位点(5ʹSS)和分支点序列分别被 U6 和 U2snRNA 特异性识别。除了催化金属离子的配位作用外,Prp8 催化空腔中的 RNA 元件大多为催化作用做好了准备。催化潜伏阶段由包围 BPS 的 SF3B 复合物以及掩盖 5ʹ外显子- 5ʹSS 连接的剪接因子 Cwc24 和 Prp11 维持。这个结构与前面确定的结构一起勾勒出了前 mRNA 剪接反应的分子框架。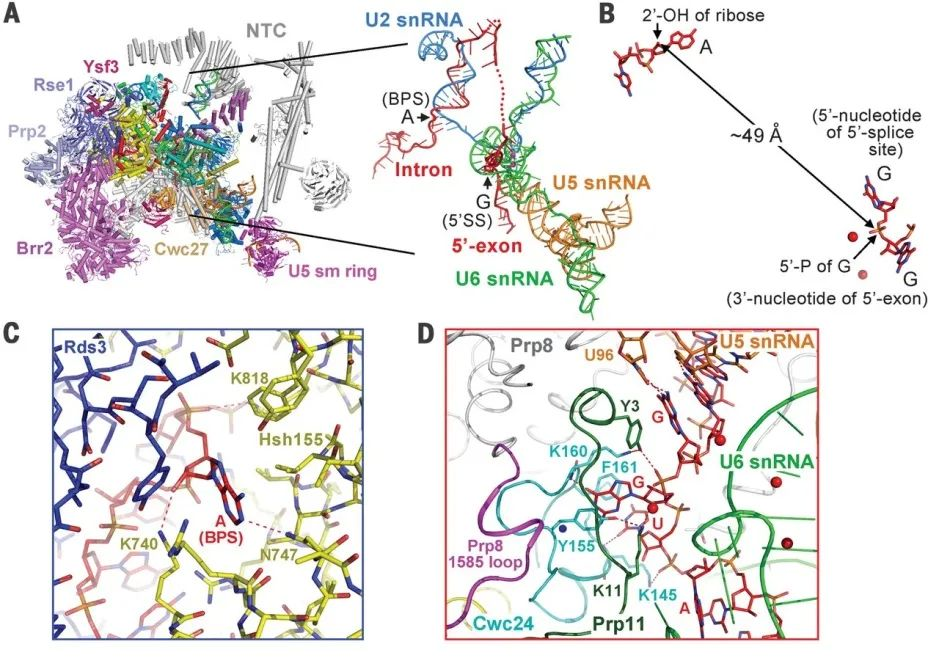
Bact 复合物的催化潜伏期(图片来源:Science)
3. Structure of a yeast catalytic step I spliceosome at 3.4 Å resolution
(Science 26 Aug 2016)
由剪接体执行的前信使 RNA 剪接作用中,每个循环包括两个逐步发生的酯交换反应,这两个反应移除了一个内含子并连接了两个外显子。
这篇文章研究了来自酿酒酵母的催化步骤 I 剪接体(称为 C 复合物)的原子结构,这是由平均分辨率为 3.4 埃的冷冻电子显微镜测定的。这篇研究与上一篇研究论文于同日发表在 Science 杂志。
在结构中,分支点序列(BPS)中不变腺嘌呤核苷酸的 2’-OH 与 5’剪接位点(5’SS)5’端的磷酸共价连接,形成内含子套索。游离的 5’外显子仍然锚定在 U5 小核 RNA(SnRNA)的环 I 上,内含子的 5’SS 和 BPS 分别与保守的 U6 和 U2 snRNA 序列形成双链。这些 RNA 元件在 Prp8 催化空腔的特异性放置由 15 个蛋白质组分稳定,包括 Snu114 和剪接因子 Cwc21、Cwc22、Cwc25 和 Yju2。这些特征揭示了第一步反应后剪接体的构象,并预测了执行第二步酯交换反应所需的结构变化。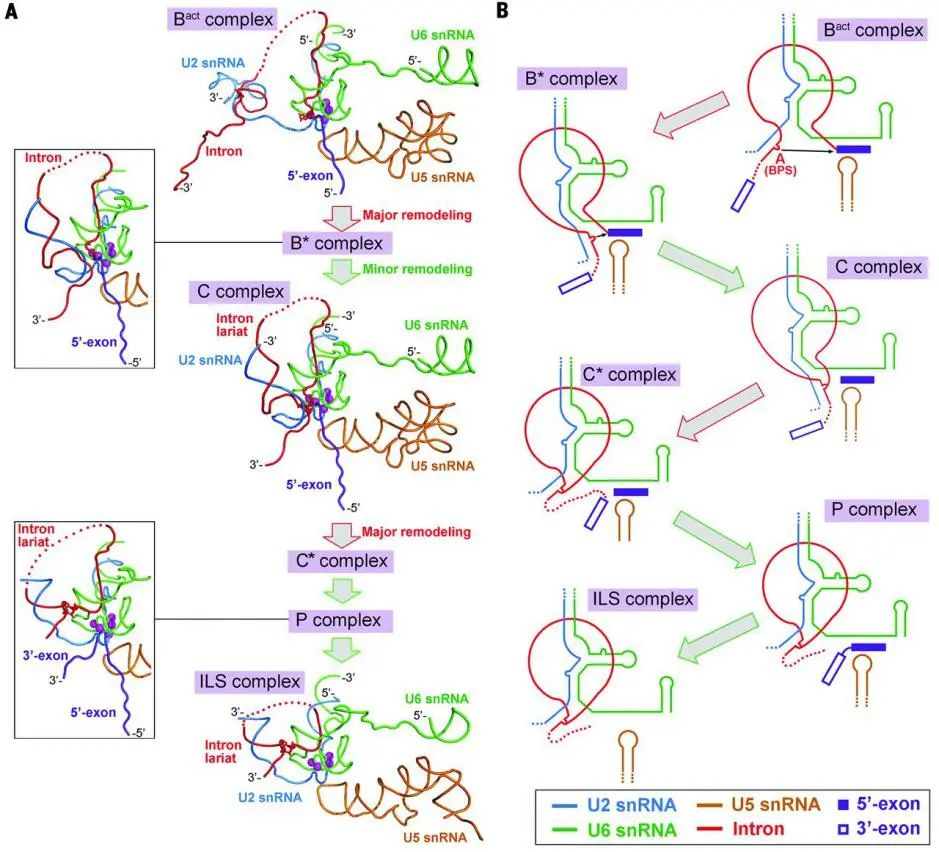
RNA 构象水平上前 mRNA 剪接的工作模式(图片来源:Science)
4. Structure of a yeast step II catalytically activated spliceosome
(Science 13 Jan 2017)
(图片来源:Science)
前体信使 RNA(Pre-mRNA)剪接的每个循环包括两个逐步发生的反应,首先释放 5’外显子并产生内含子套索- 3′外显子,然后连接两个外显子并释放内含子套索。第二个反应由步骤 II 催化激活的剪接体 (称为 undefined 复合物) 执行。
这篇文章中,作者探究了来自酿酒酵母的 C~_复合物的冷冻电子显微镜结构,其平均分辨率为 4.0 埃。与之前的剪接体复合体(C 复合体)相比,套索连接移位了 15~20 埃,为传入的 3’-外显子序列腾出了空间。
步骤 I 剪接因子 Cwc25 和 Yju2 已从活性位点解离。Prp8 的两个催化基序 (核酸酶类 H 结构域的 158 环,和 β 指),以及步骤 II 剪接因子 Prp17 和 Prp18 和其他周围蛋白,潜在促进第二次酯交换。这些结构特征加上其他剪接体复合物的研究报道,完善了关于剪接循环的机制图。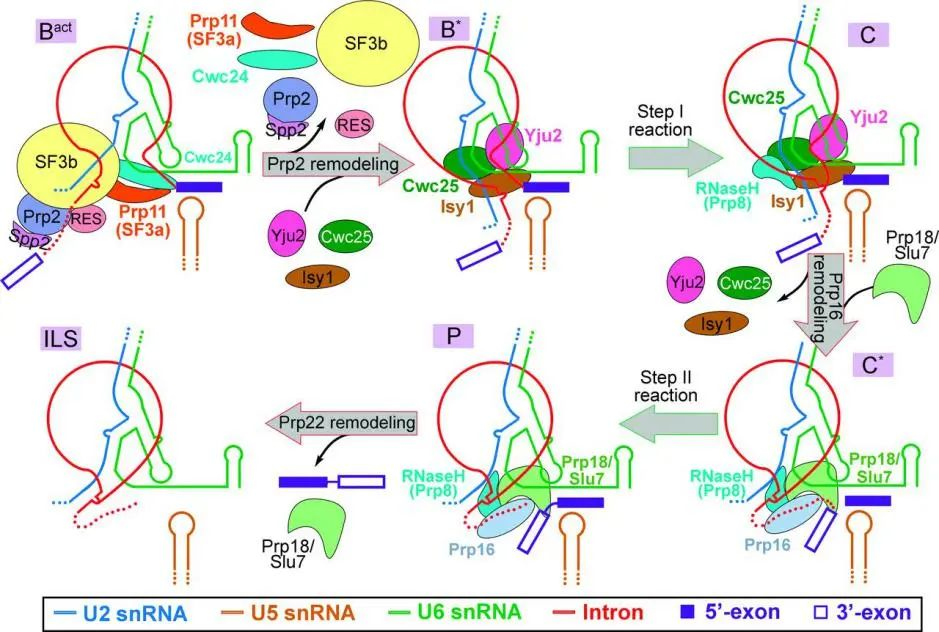
Pre-mRNA 剪接的机制模型(图片来源:Science)
5. Structure of an Intron Lariat Spliceosome from Saccharomyces cerevisiae
(Cell 14 September 2017)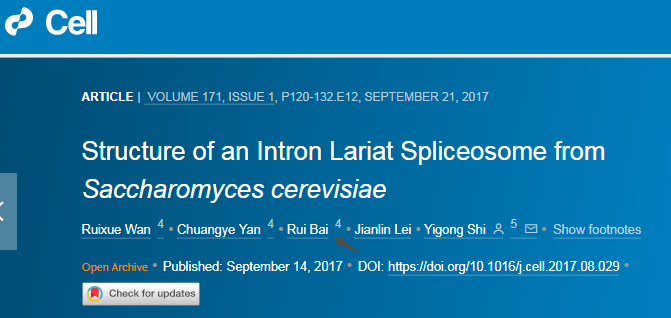
(图片来源:Cell)
内含子套索剪接体(ILS)的解体标志着剪接循环的结束。
这篇文章报道了酿酒酵母 ILS 复合物的低温电子显微镜结构,其平均分辨率为 3.5 埃。内含子套索仍然结合在剪接体中,而连接的外显子已经解离。步骤 II 剪接因子 Prp17 和 Prp18 以及稳定与 U5 小核 RNA(SnRNA)环 I 结合的 5’外显子的 Cwc21 和 Cwc22 已经从活性位点组装中释放。
Deah 家族 ATP 酶/解旋酶 Prp43 与剪接体外围的 Syf1 结合,其 RNA 结合位点接近 U6snRNA 的 3’端。Ntr1 /Spp382 的 C-末端结构域与 GTPase Snu114 结合,而 Ntr2 与 Ntr1 的超螺旋结构域相互作用时锚定在 Prp8 上。这些结构特征揭示了 ILS 复合物的一种可靠的拆解机制。
6. Structure of the Post-catalytic Spliceosome from Saccharomyces cerevisiae
(Cell 14 December 2017)

(图片来源:Cell)
剪接体从前 mRNA 中去除内含子会导致催化后剪接体中两个外显子的连接(称为 P 复合体)。
这篇文章中报道了来自酿酒酵母的 P 复合体的低温 EM 结构,平均分辨率为 3.6 埃。首次展示了 pre-mRNA 中 3' 剪接位点的识别状态,该结构为回答 RNA 剪接反应过程中 pre-mRNA 中的 3' 剪接位点如何被识别,第二步转酯反应如何发生以及成熟的 mRNA 如何被释放等关键问题提供了重要的结构信息。
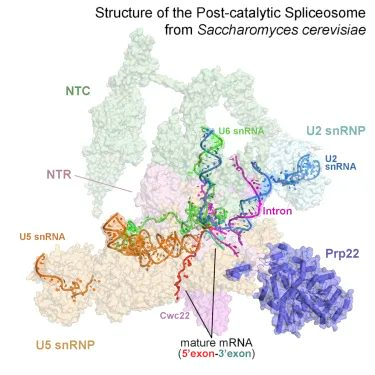
(图片来源:Cell)
7. Structures of the Catalytically Activated Yeast Spliceosome Reveal the Mechanism of Branching
(Cell 4 April 2018)

(图片来源:Cell)
前 mRNA 剪接是由剪接体执行的。催化活化配合物(B-∗)的结构表征是理解支化反应的关键。
在本研究中,作者从酿酒酵母的两个不同的 Pre-∗上组装了 B-∗复合物,并测定了四个不同的 B-RNA 复合物的冷冻-EM 结构,总分辨率为 2.9 - 3.8 埃。
在 3 个不含步骤 I 剪接因子 Yju2 和 Cwc25 的 B∗复合物中,U2 小核糖核酸和分支点序列之间的双链与 5’-剪接位点(5’SS)分开。Yju2 被招募到活性中心使 U2 /BPS 双链进入 5’SS 附近,而 BPS 亲核基团位于距离催化金属 M2 4 埃的位置。这一分析揭示了 Yju2 和 Cwc25 在分支中的作用机制。不同前 mRNA 上的这些结构揭示了剪接体在主要功能状态下的底物特异性构象。
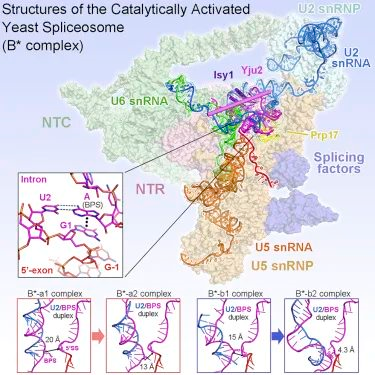
(图片来源:Cell)
8. Structures of the fully assembled Saccharomyces cerevisiae spliceosome before activation
(Science 29 Jun 2018)
(图片来源:Science)
前催化剪接体(B 复合体)由前 B 复合体变化而来。这篇文章研究了酿酒酵母前 B 和 B 复合物的冷冻电子显微镜结构,其平均分辨率分别为 3.3 到 4.6 埃和 3.9 埃。
这是一个非常重大的突破,揭示了世界上最大、最复杂的剪接体结构,也是施一公实验室第一次捕获并解析了一个全新的剪接体状态,审稿人更是将该结构评价为史上最重要、最振奋人心的剪接体结构之一,在领域内有极大的反响。
在 Pre-B 复合体中,5’剪接位点 (5’sS) 和 U1 小核 RNA(SnRNA)之间的双链被 Yhc1、Luc7 和 Sm 环识别。
在 B 复合体中,U1 小核糖核蛋白被解离,5’-外显子- 5’sS 序列移位到 U6snRNA 附近,3 个 B-特异性蛋白可能定位前体信使 RNA。
在这两个复合物中,U6 snRNA 锚定在 U5 snRNA 的环 I 上,分支点序列和 U2 snRNA 之间的双链被 SF3B 复合物识别。Pre-B 复合体的结构阐明填补了剪接体对 Pre-mRNA 剪接的机制理解中的重要知识空白,揭示了酵母剪接体的组装和激活机制。
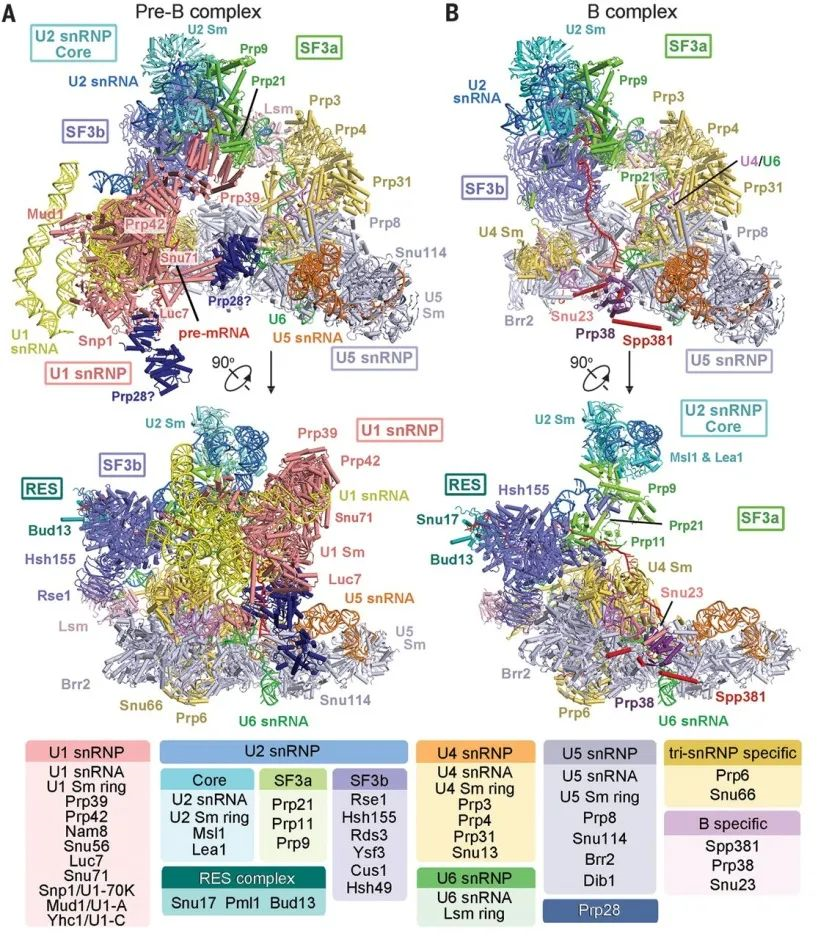
酿酒酵母 Pre-B 和 B 复合体的冷冻电镜结构(图片来源:Science)
9. Mechanism of spliceosome remodeling by the ATPase/helicase Prp2 and its coactivator Spp2
(Science,26 Nov 2020)
通过阻滞方法获得了高分辨率的原位酿酒酵母剪接体(Bact complex)的电镜结构,解析了 ATP 酶/解旋酶 Prp2 及其共激活剂 Spp2 重塑剪接体的机制,是国际上第一篇剪接体重塑机制的结构研究。
白蕊、施一公等揭示剪接体激活过程中结构重塑的分子机理(图片来源:Science)
由保守的 ATP 酶/解旋酶(包括 Prp2)执行的剪接体重塑可以实现前体 mRNA 的剪接。然而目前对 ATP 酶/解旋酶功能的结构基础仍然知之甚少。
在作者报道的 Bact 复合体中观察到了未在结构研究中(如酵母或人类 Bact)观察到的 RNA 解旋酶/ATP 酶 Prp2、其激活因子 G 蛋白 Spp2 及与 Prp2 结合识别的底物 pre-mRNA。
作者进而结合生化实验,阐明了剪接体 Bact complex 重塑的发动机 Prp2 如何被募集到剪接体上、Spp2 如何稳定 Prp2 以及 Prp2 为何具有使底物 pre-mRNA 向其 3’端单向移动的方向性等关键科学问题。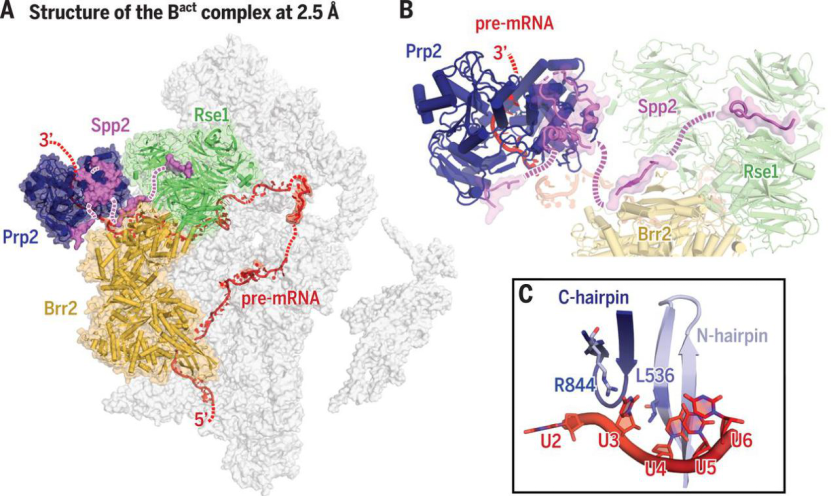
ATP 酶/解旋酶 Prp2 在活化剪接体中的作用机制(图片来源:Science)
10. Structure of the Activated Human Minor Spliceosome
(Science,28 Jan 2021)
(图片来源:Science)
稀有内含子如 U12 被次要剪接体去除。与主剪接体相比,次要剪接体的组成、组装、功能状态、活化、调控和结构一直是个谜。
在这项研究中,白蕊及其同事在 U12 型内含子(仅占人类基因组 1%)剪接实验中组装了活化的人源次要剪接体,并首次报道了人源次要剪接体高分辨率(2.9 Å)的三维结构,解析了在剪接反应中的关键构象——激活态次要剪接体(次要 Bact 复合物)的结构及对稀有内含子(U12 依赖型内含子)的识别剪接机理。
主要 Bact 复合物的 13 种蛋白质,包括 5 种 NTC 和 NTR 组分,在次要 Bact 复合物的结构中不存在。
作者在活化的次要剪接体中发现五种发挥重要作用新蛋白 (ARMC7,CRIPT,RBM48,SCNM1 和 PPIL2)。5 个新鉴定的蛋白质稳定了催化中心的构象: 锌指蛋白 SCNM1 功能上模拟主要剪接体的 SF3a 复合物,RBM48-ARMC7 复合物结合 U6atac snRNA 5’末端的 γ-单甲基磷酸帽,U-box 蛋白质 PPIL2 协调 U5 snRNA 的环 I 并稳定 U5 小核核糖核蛋白(snRNP),CRIPT 稳定 U12 snRNP。
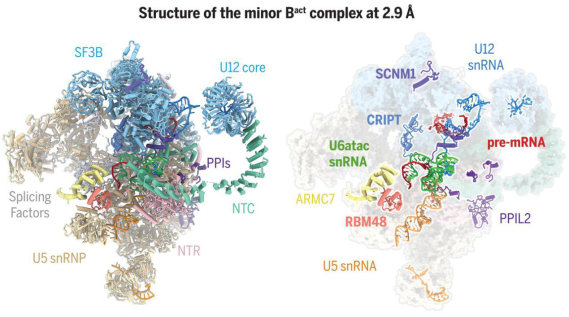
(图片来源:Science)
优秀的一个个成果背后,是白蕊坚定的信念、坚持和努力,读博期间,她也曾因病痛低落迷茫至想要退学,也曾因为被竞争对手抢发文章而难过到彻夜难眠,这些都是科研道路上的万千难关,她都一一克服。
白蕊说:「我做科研最大的动力是,想治疗某一种疾病。」期待在不久的将来,她的科研成果可以造福更多的患者。











