外源基因在大肠杆菌中的诱导表达和降解物阻遏作用
互联网
| 外源基因在大肠杆菌中的诱导表达和降解物阻遏作用 |
| 【实验目的】 |
|
1.了解外源基因在原核细胞中表达的基础理论。 |
| 【实验原理】 |
|
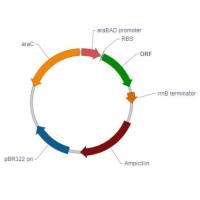
1.外源基因在原核细胞中的表达 蛋白质通常是研究的最终目标,因此蛋白质的表达在基因工程中占有非常重要的地位。常用的表达系统有原核细胞和真核细胞。 原核细胞表达系统主要使用大肠杆菌,真核细胞表达系统主要有酵母细胞、哺乳动物细胞和昆虫细胞。这些表达系统各有优缺点,应根据实验目的和实验室条件加以选择。本实验主要介绍以大肠杆菌为代表的原核细胞表达系统。 (1)大肠杆菌表达系统的特点: 而在真核基因上则缺乏该序列。因此,一些商品化原核表达载体上设计有SD序列,以方便真核基因的表达。 原核细胞没有mRNA转录后加工的能力。因此,在原核细胞中表达真核基因时,应使用cDNA为目的基因。 常用的原核启动子有:由异丙基-D-硫代半乳糖苷(IPTG)诱导的lac启动子,由3-吲哚乙酸(IAA)诱导的trp启动子,由温度诱导的PL和PR启动子等。噬菌体 T7 RNA聚合酶启动子是一个很强的启动子,近年来在原核表达中得到广泛应用。 非融合蛋白的一级结构和天然蛋白质相同,是一些体内应用基因工程产品的必要条件。但是非融合蛋白在原核细胞内不稳定,易被降解,而且不易纯化。 常用的信号肽有ompT、phoA、pelB等,在表达的蛋白进入细胞周间质时,信号肽被蛋白酶水解,产生游离的表达产物。因此,分泌型表达可以保护外源蛋白不被细胞内的蛋白酶降解,增加表达产物的稳定性,同时,表达蛋白的生物活性较好,易于纯化,但是,表达量往往比较低。
操纵子是原核细胞基因表达的协调单位。通常由两个以上功能相关的结构基因以及一些调节序列(如启动子序列、操纵序列等)组成。乳糖操纵子由三个结构基因Z、Y、A和操纵序列、启动子、CAP结合位点等调节序列组成。 乳糖操纵子的调节包括乳糖(或IPTG)的诱导效应和葡萄糖的降解物阻遏效应。 (1)乳糖操纵子的诱导表达 当没有乳糖存在时,调节基因lacI表达,转录的mRNA翻译成阻遏蛋白。阻遏蛋白与操纵序列lacO结合,阻碍了结合在旁边启动子的RNA聚合酶向前移动,使目的基因(本实验中目的基因为绿色荧光蛋白基因)不能转录,也就不能翻译出目的蛋白。也就是说,当没有乳糖存在时,乳糖操纵子处于阻碍状态。 当有乳糖存在时,乳糖转化为异乳糖,异乳糖作为诱导物与阻遏蛋白结合,使阻遏蛋白的构象发生改变,而不能结合到操纵序列上,RNA聚合酶可以从启动子向3′端移动,于是,结构基因可以转录出mRNA,然后翻译出蛋白质。也就是说,当有乳糖存在时,乳糖操纵子被诱导(如图3)。
乳糖操纵子的诱导物是异乳糖。IPTG是异乳糖的结构类似物。由于IPTG不会被分解,它的诱导作用是持久的。
当CAP与cAMP结合后,就可结合到CAP结合位点上,促进转录。葡萄糖降解物能抑制腺苷酸环化酶的活性,并活化磷酸二酯酶的活性,从而降低cAMP的浓度,抑制转录。 |
| 【试剂与器材】 |
|
(一)试剂 (二)器材 (三)菌株 |
| 【操作方法】 |
|
1.分别挑取工程菌BL21(DE3)pET-32a和BL21(DE3)p32aGFPuv的单菌落接种到5mLLB液体培养基(含氨苄,终浓度为100μg/mL,以下同)。 2.于37℃、250r/min培养过夜(12h-14h)至对数生长期。 3.取三支灭菌试管,各加入5mL LB液体培养基(含氨苄),分别编号为1#,2#,3#,另外取装于500mL三角瓶中的150mL LB液体培养基(含氨苄),编号6#, 全部按1:50的比例接种。1# 接种100μL BL21(DE3)pET-32a,2# 接种100μL BL21(DE3)p32aGFPuv,3# 接种100μL BL21(DE3)p32aGFPuv,6# 接种3mL BL21(DE3)p32aGFPuv。 4.于37℃、250r/min培养约2h-3h。 5.诱导处理:1#、2#不需处理;3#加入20%葡萄糖100μL至终浓度为0.4%,加入5μL 100mmol/L IPTG至终浓度为0.1mmol/L;6#加入100mmol/L IPTG约150 μL至终浓度为0.1mmol/L。 6.于21-25℃、250r/min培养约10h-12h或过夜。 7.收菌 (注意采集标本并编号): ①从6#三角瓶培养的150mL菌液中取出5mL置于一支试管中(编号为4#)。 ②分别从1#,2# ,3#培养物各取100μL于EP管用于SDS-PAGE。 ③将1#,2#,3#,4#试管中的菌液分别收集到4个1.5mLEP离心管中,编上相应编号。1#,2#,3#离心弃上清;4#上清收集到另一个EP管,编号为5#。 ④6#三角瓶中剩余菌液用 50mL离心管于5000r/min,4℃,离心10min,弃上清收集菌体。 8.于紫外灯下观察1# -5#管收集的菌体或上清液,观察哪支管有荧光(荧光强弱),记录观察到的现象,并拍照。 9.6#收集的全部菌体用50mL超声平衡缓冲液重悬(50mmol/L Tris-HCl,500mmol/L NaCl pH7.0)。 10.超声波破菌或高压破菌,然后用50mL离心管于8000r/min,4℃,离心40min,取上清(弃沉淀)。上清可保存于-20℃冰箱,作下一步亲和层析实验用。 超声操作:冰浴下进行,功率为400W,工作4s,间隙4s,为一次,99次为一周期。共处理六周期。 |
| 【注意事项与提示】 |
|
1.本实验的目的是比较重组DNA在大肠杆菌中表达时是否受IPTG和葡萄糖存在的影响,所以1#、2#管的细菌在25-28℃培养时不加IPTG和葡萄糖,3#管除加IPTG外还加入葡萄糖,糖的浓度要大于0.2%以上(本实验采用0.4%)。 4#三角瓶细菌仅加IPTG 也仅指这次实验所用的重组DNA菌体而言。有的重组DNA菌体在其表达时除需加IPTG外仍需加入少量的葡萄糖为碳源作诱导。在转管培养及收菌时要注意各管编号要相应对好,不能混乱。 2.不同的重组DNA在不同的宿主菌中蛋白的表达量往往受到IPTG浓度和温度的影响,最好采用不同温度或不同浓度的IPTG来诱导,观察哪种温度或哪种浓度条件下其蛋白表达量最大。在科研中一般都要求这样做。 3.由于本实验所表达的目的蛋白带有绿色荧光蛋白,其在大肠杆菌中有很强的荧光。离心后收集的菌体在紫外线的照射下都可见黄绿色荧光,所以把1#、2#、3#、4#沉淀菌体放在紫外灯下观察其是否有荧光及其荧光的强弱,就可判断其是否有表达及其所表达的强度。 |
| 【实验安排】 |
|
1.第一天晚上9时左右活化种子菌。 |
| 【实验报告要求与思考题】 |
|
1.对紫外灯下观察到的结果作出解释。 |