T细胞抗原受体TCR多样性的产生机制
互联网
TCR多样性(个体水平)最终可达到1015~1018,形成容量庞大的TCR库,赋予个体几乎是无限的抗原识别和应答能力,保证个体在多变环境中能和外来抗原(病原体)发生有效的免疫应答。下面简述TCR多样性的产生机制。
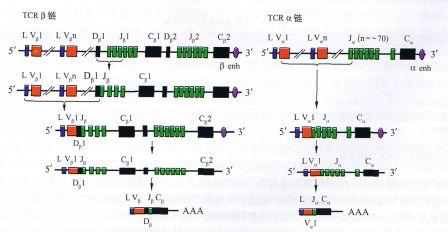
(1)多个胚系基因片段的组合多样性:TCR的胚系基因由多个分隔的基因片段组成(表2―3),它们在基因重排过程中的随机组合为多样性的产生提供了遗传学基础。
(2)VJ和VDJ连接(重排)多样性:基因重排系一随机过程,不同的T细胞克隆经基因重排、发生不同基因片段的连接,产生特定的Vp(VDJ)基因和Va(VJ)基因,表达特异性的TCR,这是TCR多样性产生的主要机制。
(3)连接机动性(junctionalflexibility):也称连接不精确性(junctionalimprision)。V区基因重排时,特定的V、D或J片段在发生DJ、V―DJ或VJ连接时,由于读码框的机动性或不精确性,在V―D-J连接处可发生一定的变异(偏移)导致核苷酸缺失,引起核苷酸序列、密码子的改变,最后导致V区氨基酸组成的变化,从而改变TCR的特异性,增加了TCR的多样性。
(4)N区核苷酸插入:N区核苷酸插入(N-regionnucleotideaddition)发生在V区基因重排过程中。N区核苷酸片段(non-germlinegeneencodednucleotides)并不存在于TCR的胚系基因中。
V区基因重排时,在末端脱氧核苷酸转移酶(TdT)作用下,N区核苷酸片段可随机加在Vα的VJ连接处及Vβ的VDJ的D片段的两侧。N区核苷酸片段最多可由6个核苷酸组成,且富含G(鸟嘌呤)和C(胞嘧啶),防止形成终止密码(UAA、UGA和UAG或TAG)。N区核苷酸插入也发生在CDR3区。
(5)α链、β链的组合多样性:TCR的V区由Vα和Vβ组成,它们的随机配对和组合参与构成TCR的多样性。
上述五种机制赋予个体产生容量庞大的TCR库,并通过TCR的V区体现出来,其中以CDR3的变异最大,决定了TCR的特异性(克隆水平)和多样性(个体水平)。