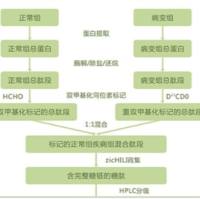
蛋白质组学在植物科学研究中的应用
互联网
1 植物群体遗传蛋白质组学
1.l 遗传多样性蛋白质研究
基于基因组学的一些遗传标记,如RAPD(Random Amplified Polymorphic DNA)、RFLP(Restriction Fragment Length Polymorphism)、SSR(Simple Sequence Repeat)、ISSR(Inter-Simple Sequence Repeat)等,已经广泛地应用于植物遗传研究中。
与基因组学的遗传标记相比,由于蛋白质组学的研究对象是基因表达的产物,是介于基因型和表型之间的特性,因而蛋白质组学标记是联系基因多样性和表型多样性的纽带,具有独特的意义。
通过蛋白质组比较来检测遗传多样性的变化已有许多成功的尝试。Barrenche等(1996年)比较了6个欧洲国家的23种橡树,分析了幼苗的总蛋白质,共得到530种蛋白质,其中101个具有多态性。
实验结果显示种内和种间的距离非常接近,并且证实无梗花栎(Quercus petraea)和夏栎(Quercus robar)两个种的遗传分化水平很低。Picard等(1997年)利用2D-PAGE分析了亲缘关系很近的硬粒小麦不同株系的遗传多样性,发现品系间的多态性很低并且7个蛋白可以用于基因型的鉴定。
David等(1997等)也利用2D-PAGE技术比较了栽培于不同环境下但起源于同一种群的小麦,结果所有的种群都与原种群有差别,David等认为,这不是由随机漂移引起,而是由适应其各自的气候条件而形成。
1.2 突变体的蛋白质组学研究
突变体研究是植物遗传学的重要研究手段之一,应用蛋白质组学的方法对基因突变引起的蛋白质表达变化进行研究可以揭示一些植物生理生态过程的机制。具体做法通常是对在相同条件下栽培的突变体及野生型植物的2D-PAGE图谱进行比较,受到影响的蛋白质通过质谱法或Edman测序法进行鉴定,为研究表型突变背后的生化过程提供有价值的信息。
Santoni等(1994年)对模式植物拟南芥发育突变体的总蛋白质进行了分析,结果显示突变体具有与野生型植物不同的独特的2D-PAGE图谱,并且得到了与下胚轴长度有关的一个肌动蛋白的同源异构体。
Herbik等(1996年)分析了野生型和缺铁突变体番茄(Lycopersicon esculentum)的蛋白质2D-PAGE图谱,鉴定了参与无氧代谢和胁迫防御的几种酶,如甘油醛-3-磷酸脱氢酶、甲酸脱氢酶、抗坏血酸过氧化物酶、超氧化物歧化酶、质体蓝素等,并分析了这些酶在获得铁的过程中的功能。
von Wiren等(1997年)比较了野生型和铁摄取缺陷型突变体玉米的蛋白质2D-PAGE图谱,确定了4个与铁离子跨膜运输有关的多肽。
Komatsu等(1999年)比较了水稻绿苗和白化苗的蛋白质2D-PAGE图谱,发现了在绿苗中参与光合作用的蛋白质,而白化苗中仅有此蛋白质前体。而抗坏血酸过氧化物酶只在白化苗中存在,说明抗氧化酶在白化苗中起细胞保护功能。
当已知突变的基因时,可用蛋白质组学技术研究受此基因控制的有关信息。玉米中的Opaque 2(O2)基因编码一个属于亮氨酸拉链家族的转录因子,这个转录因子对蛋白质的表达有多种效应。
Damerval和Le Guillonx(1998年)将野生型与O2基因突变体的蛋白质2D-PAGE图谱进行比较,鉴定出属于各种代谢途径的一些酶,说明O2基因是玉米代谢中联系多种代谢途径的调节基因。这些结果表明,单一位点的突变可以引起蛋白质表达的多种效应,而用蛋白质组学技术可以看到这些效应。
2 植物环境信号应答和适应机制蛋白质组学
2.1 非生物环境因子蛋白质组学研究
在植物的生存环境中,一些非生物因子胁迫,如干旱、盐渍、寒害、臭氧、缺氧、机械损伤等,对植物的生长发育和生存都会产生严重影响。这些胁迫可以引起大量的蛋白质在种类和表达量上的变化,而蛋白质组学研究可以使我们更好地了解非生物胁迫的伤害机制以及植物对非生物环境的适应机制。
Salekdeh等(2002年)研究两个水稻品种(Oryza sativa L. cv CT9993 和 cv IR62266)干旱胁迫下以及恢复灌溉后的蛋白质组。分析叶提取物电泳胶上的1000多个蛋白点,发现有42个蛋白点的丰度在干旱胁迫状态下变化明显,其中27个点在两个品种中显示了不同的反应方式。
恢复正常灌溉10天以后,所有蛋白的丰度完全或是在很大程度上恢复成与对照一样。Costa等(1998年)发现海岸松(Pinus pinaster)中38个受干旱影响的蛋白质,其中24个由干旱诱导,并且不同基因型对干旱胁迫的反应差别很大。
Ramani和APte(1997年)用放射性同位素自显影双向电泳法研究水稻幼苗盐胁迫下多基因的瞬时表达表明,至少有35个蛋白质被盐胁迫诱导和17个蛋白质被抑制,包括20个在这之前未曾报道的低丰度蛋白。这些发现对寻找渗透压应答新基因,尤其是那些在水稻盐耐性获得中起瞬时调节作用的基因十分重要。
Agrawal等(2002年)用2D-PAGE、氨基酸测序和免疫杂交法,首次检测了臭氧对水稻幼苗蛋白的影响。臭氧对叶片强烈的可见坏死伤害和随之而来的抗坏血酸过氧化物酶蛋白的增加,反映在二维电泳胶上蛋白质点分布的变化。
在被检测的具有可重复结果的56个蛋白中,52个蛋白点随控制条件的不同发生的变化可以通过肉眼判定。检测的56个蛋白中,6个蛋白是N-端阻断,14个蛋白的序列无法测定,36个蛋白的N-端序列和一个蛋白的内部序列被测定。研究发现臭氧造成叶片光合蛋白的剧烈减少(包括核酮糖-1,5-二磷酸羧化酶/加氧酶)和各种防御、胁迫相关蛋白的表达。
Shen等(2003年)应用蛋白质组学方法研究水稻叶鞘伤害反应相关蛋白,首次揭示了水稻叶鞘伤害信号应答过程中蛋白质的变化。比较伤害前后蛋白质表达谱,发现伤害后至少有10个蛋白被诱导或上调,19个蛋白被抑制或表达量下降。
通过N-端或内部氨基酸测序,分析了其中的14个蛋白,鉴定了9个蛋白的功能,其它蛋白由于N-端阻断,无法得到氨基酸序列信息。此外,还通过MALDI-TOF-MS测定了11种蛋白质,并与水稻数据库相吻合。
在基因功能被确认的蛋白质中,表达量下降的蛋白有2个钙网蛋白、组蛋白H1和血红蛋白和一种假定的过氧化物酶;表达量增加的蛋白包括胰蛋白酶抑制因子(BBT1)、两种假定的蛋白激酶受体类似物、钙调素相关蛋白、核酮糖-l,5-二磷酸羧化酶/加氧酶小亚基、2个甘露糖结合外源凝集素。
其中,4种蛋白质已被证实为与伤害反应直接作用的蛋白质。
Chang等(2000年)对玉米进行缺氧和低氧胁迫研究,发现低氧处理的效应不仅仅是缺氧胁迫诱导的糖酵解酶的增加。通过质谱法共鉴定了46个蛋白质,均为在植物中首次得到鉴定。
2.2 生物环境因子蛋白质组研究
植物的生长发育不仅与非生物因子密切相关,还受到生物因子如动物、植物、微生物等的极大影响。例如,当植物受到竞争、动物取食、微生物共生或寄生、病菌侵害时,植物将改变体内蛋白质的表达和酶类的活性等来完成这些信号的感应、传递以及生物学效应的实现。
因此,通过对蛋白质的研究有助于人们更好地了解生物之间的相互作用机制。目前关于非生物因子对植物影响的蛋白质组学研究主要集中在植物与根瘤菌以及植物与菌根真菌的共生关系上。
众所周知,根瘤菌与植物相互识别后,进入植物细胞内变成具有因氮能力的类菌体。类菌体周隙(PS,Peribacteroid space)是类菌体周膜(Peribacteroid membrance,PBM)与细菌质膜之间的间隙,是共生体成员之间交换代谢产物的媒介。
Saalbach等(2002年)用蛋白质组分析的方法,鉴定了PBM与PS中的蛋白。结果表明PS甚至PBM的制备物中含有大量的类菌体蛋白。有趣的是,除了一些PS/PBM蛋白,还有许多内膜蛋白,包括V-ATPase,BIP和一个完整的COPI-coated vesicles的膜蛋白存在于PRM中,这证明了PBM是由宿主细胞的内膜系统产生的。
Wienkoop和Saalbach(2003年)选取豆科模式植物日本百脉根(Lotus japonicus),用蛋白质组学的手段研究PBM的蛋白质组。通过纳米液相色谱分离多肽,然后用串联质谱(MS/MS)进行分析。
检索非丰度蛋白数据库和通过串联质谱得到的绿色植物表达序列标签数据库,鉴定了大约94个蛋白,远远多于迄今为止所报道的PBM蛋白的数目。
特别是一些膜蛋白得到检测,如糖和硫酸盐转运子、内膜联合蛋白(如GIP结合蛋白)和囊泡受体、信号相关蛋白(如受体激酶、Calmodulin、14-3-3蛋白和病原体应答蛋白包括HIR蛋白)。通过非变性凝胶电泳分析了两个特征蛋白复合物。结果鉴定了PBM中参与特定生理过程的蛋白和结瘤特异表达序列标签数据库(EST)中的PBM蛋白质组。
Bestel-Corre等(2002年)用双向凝胶电泳和银染分析接种灌木菌根真菌(Glomus mosseae)或根瘤菌(Sinorhizobiurn meliloti)的模式植物苜蓿不同时期的根蛋白质组。MALDI-TOF-MS分析胰蛋白酶消化的蛋白质组,在结瘤的根中鉴定到了苜蓿的一个豆血红蛋白。内部测序、四极质谱分析和数据搜寻证明了先前预测的由菌共生体诱发表达的蛋白。
2.3 植物激素蛋白质组学研究
激素在植物一生中起着重要的调控作用,研究植物激素的信号传导和作用机理是蛋白质组学的重要组成部分之一。
Moons等(1997年)鉴定了水稻根中3个受ABA诱导的蛋白质,其氨基酸序列测定确定其中2个属于胚胎后期丰富蛋白(LEA)的2组和3组,第三个未知。
用ABA处理水稻根并提取其mRNA,构建cDNA文库,然后用由氨基酸序列推导出的寡核苷酸做探针进行筛选,分离到了相关的cDNA,但在cDNA数据库中找不到与之相同的序列。通过在基因组DNA文库中筛选及免疫杂交分析,表明这是一个新的基因家族,编码高度亲水具有双重结构域的蛋白质,并被ABA诱导在不同组织中表达。
Rey等(1998年)在马铃薯的叶绿体中发现了一个受干旱诱导的蛋白质,没有已知的蛋白质与之相同。利用N-端序列制备的血清在叶子cDNA表达文库中筛选,分离到了新的具有典型硫氧还蛋白特征的cDNA序列,并被硫氧还蛋白活性的生化实验所证实。
蛋白质组学技术不仅可鉴定早就已知的典型的受环境胁迫诱导的蛋白质,如LEA蛋白质(Riccardi等,1998年),脱水素(Dehydrin)(Moons等,1997年),还鉴定了其它一些蛋白质。
如对胁迫引起的损害起保护作用的蛋白酶抑制剂、热激蛋白HSP和与氧化胁迫有关的酶、参与糖酵解、木质素合成的酶等(Costa等,1998年;Riccardi等,1998年;Rey等,1998年;Pruvot等,1996年)。用赤霉素和茉莉酸处理水稻的蛋白质组变化也进行了研究。
Shen和Komatsu(2003年)将水稻叶鞘用5μmol·L-1赤霉素处理不同时间后的蛋白经2D-PAGE分离和计算机图像分析,看到33个蛋白发生变化,其中21个蛋白点表达增强,12个蛋白表达减弱,说明赤霉素处理水稻叶鞘起码有30多个基因的产物与之相关。
对其中的钙网蛋白(Calreticulin)进行了深入分析,发现它有2个不同等电点(pI)蛋白点,随赤霉素处理时间增加,pI4.0的蛋白点逐渐消失,而pI4.1蛋白点浓度则逐渐增加。由此说明钙网蛋白在赤霉素信号传递调节叶鞘伸长中是一个重要组分(Shen等,2003年)。
Rakwal和Komatsu(2000年)用外源茉莉酸(Jasmonic acid)处理水稻的幼苗组织,通过2D-PAGE分析发现在水稻的茎和叶中诱导了新蛋白质。对蛋白质点进行N-端和内部测序及免疫杂交分析,发现茎中有28kD的蛋白酶抑制剂(BBPIN)和酸性的与病理有关的17kD蛋白质(PR-1)。
免疫杂交分析表明茉莉酸处理后这些蛋白质的表达具有组织特异性和发育阶段特异性,说明外源茉莉酸处理可以引起与植物自我防御机制有关的基因在茎、叶组织中的特异性表达。
3 植物组织器官蛋白质组学
对于植物来说,蛋白质组学上的差异不但存在于不同基因型以及同一基因型的不同植株之间,也存在于同一植抹的不同组织和器官之间。在植物的发育过程中,不同组织和器官在功能上的分化,也表现在不同器官蛋白质的组成和数量的差异上,蛋白质组学的研究有助于我们对植物发育过程机制的理解。
关于植物组织和器官的蛋白质组学研究已经有很多报道。Tsugita等(1994年)用2D-PAGE分离了水稻根、茎、叶、种子、芽、种皮及愈伤组织等部位的蛋白质,总共得到4892个蛋白点,其中3%的蛋白质得到了鉴定。对水稻胚、胚乳、叶鞘和悬浮细胞蛋白质组学的研究也取得了进展,并且水稻的蛋白质数据库已经建立(Komatsu等, 1993年;1999年;Zhong等,1997年;Shen等,2003年)。其它组织及器官,如拟南芥的愈伤组织和花粉,也有研究涉及(Prime等,2000年;Mayfield等,2001年)。
Blee等(2001年)研究了转基因烟草(Nicotiana tabacum)细胞壁的蛋白质组。他们首先建立了转入Tcyt基因的烟草悬浮培养细胞株系。该基因可使细胞产生高水平的内源细胞分裂素,从而使该细胞株系表现出细胞聚集增加、细胞变长、细胞壁加厚5倍等特征。转化细胞壁的蛋白质组与对照烟草细胞的初生壁蛋白质组有很大差异。发现了许多初生壁中不存在的新蛋白质。已鉴定出的包括分子量为32kD的几丁质酶、34kD的过氧化物酶、65kD的多酚氧化酶和68kD的木聚糖酶,以及一些结构蛋白质。
4 植物亚细胞蛋白质组学
植物的蛋白质组学研究目前已经深入到亚细胞水平,即研究在一个细胞器内表达的蛋白质组。研究比较多的细胞器是叶绿体。据估计高等植物大约共有约21000到25000个蛋白质(Bouchez和Hoffe,1198年),叶绿体的蛋白质占其中10%~25%(van Wijk等,2000年),充分证明了叶绿体在植物细胞中的重要性。另外,关于线粒体与细胞壁的研究也有报道。
Peltier等(2000年)利用2D-PAGE、质谱及Edman N-端序列测定等方法,系统地分析了豌豆(Pisum sativum)叶绿体中类囊体的蛋白质,并在数据库中进行了搜索,鉴定了61个蛋白质,其中33个蛋白质的功能及功能结构域得到了确认。Yamaguchi和Subramanian(2000年),Yamaguchi和Subramanian(2000年)利用2D-PAGE、色谱、MS、Edman测序等多种方法鉴定了菠菜(Spinacia oleracea)叶绿体中的核糖体30S和50S亚基的蛋白质。发现菠菜的质体核糖体是由59个蛋白质组成的,其中53个与大肠杆菌有同线性,而6个是非核糖体质体特异性的蛋白质(PSRP-1到PSRP-6)。许多蛋白质表现出翻译后的修饰。PSRP蛋白质可能参与质体中特有的翻译及其调控过程,包括蛋白质通过质体50S亚基在类囊体膜上的定位和转移。
利用Blue-native凝胶电泳(Peltier等,2001年),以及MALDI-TOF和ESI-MS/MS分析,鉴定了拟南芥叶绿体中一个350kD的由10个不同的亚基组成的C1pP蛋白酶复合体,并发现了一个不属于任何已知的叶绿体基因家族的新的叶绿体蛋白。
Vener等(2001年)利用质谱技术研究了拟南芥叶绿体中类囊体膜蛋白质的磷酸化现象。研究发现,光系统Ⅱ核心中的D1、D2、CP43蛋白质位于N-端的苏氢酸(Thr)被磷酸化;外周蛋白PsbH的Thr-2被磷酸化;而成熟的光捕获蛋白LCHⅡ的Thr-3被磷酸化。Vener还研究了不同生理条件下这些蛋白质的磷酸化状态。结果表明,这些类囊体蛋白质中,没有任何一个在稳定的连续光照条件下完全磷酸化,或者在长期黑暗适应的条件下完全去磷酸化。他们还检测到在光/暗转换的条件下,PsbH的Thr-4有迅速而可逆的超磷酸化现象。D1、D2、CP43蛋白受到热激以后出现显著的去磷酸化。光合蛋白受到热激后磷酸化的变化比在光/暗转换的条件下要迅速。Vener指出,质谱法为研究复杂样品中蛋白质磷酸化的化学计量学提供了新的途径。
Peltier等(2002年)通过蛋白质组分析法与基因组预测筛选法结合,研究拟南芥叶绿体类囊体基质蛋白质组。通过双向电泳分离类囊体可溶蛋白质,再用质谱进行分析鉴定。鉴定了81个蛋白,用N-端测序对蛋白质的定位进行预测。通过实验数据修正所鉴定蛋白的基因注释,发现了一个有趣的选择性重叠。实验中还发现了大量同源基因的表达。研究表明基质蛋白质组的主要功能包括帮助折叠,催化类囊体蛋白的水解和抗氧化。鉴定的基质蛋白和它们的同源物的特性可以被应用于通过基因组预测基质蛋白质组。Schubert等(2002等)系统地描述了模式植物拟南芥类囊体基质蛋白的特性,证明类囊体基质有其自己特异的蛋白质组,并且鉴定了其中的36个蛋白。除了大量的肽基脯氨酸顺反异构酶和蛋白酶,还发现了一些新的PsbP结构域蛋白。比较模式植物拟南芥与另一个典型的高等植物菠菜的类囊体基质蛋白质组,发现二者相似性很高。作为对本实验的补充,Schubert等还从拟南芥整个基因组数据库推测基质蛋白质组,估计叶绿体类囊体基质约有80个蛋白。
位于叶绿体外膜和内膜上的参与由核编码的叶绿体蛋白质的运输蛋白质复合体得到了详尽的研究(May和Soll,1999年;Keegstra和C1ine,1999年)。膜上的疏水性蛋白质较多,用有机溶剂提取叶绿体蛋白和1D-PAGE分离,大约有5%~10%的膜蛋白,即15~20个蛋白质是疏水性的(Seigneurin-Berny等,1999年)。从绿藻(Chlamydomonas reinhardtii)中分离出一种含有酰基脂类的低密度叶绿体膜片段,类似于叶绿体内膜和类囊体膜。一些与叶绿体mRNA相结合的蛋白质非常富集,说明这些膜是叶绿体基因表达的场所(Zerges和Rochaix,1998年)。
蛋白质组学技术可以用于研究叶绿体蛋白质的翻译后修饰。这方面目前已有许多报道,包括翻译后的甲基化(对RbcS)(Grimm等,1997年),棕榈酰化(对D1)(Mattoo和Edelman,1987年)等。但是对于翻译后的修饰并没有全面、系统地开展。随着最近的2D-PAGE和质谱技术的发展和改进,这方面的研究变得更容易,这将产生许多意想不到的新发现。
蛋白质组学技术还可以对异构体基因表达及其mRNA前体的剪接、mRNA的编辑研究提供重要帮助。目前已有关于叶绿体蛋白质mRNA编辑(Sutiga和Sugiura,1996年)和剪接(Mano等,1997年)的报道。在豌豆和菠菜中已发现多基因家族(van Wijk,2000年)。这些现象当然可以通过分析mRNA或cDNA而发现,但蛋白质组学技术却是一个强大的替代或补充方法。
Heazlewood等(2003年)用等电聚焦聚丙烯酰胺凝胶电泳、B1ue-native PAGE和反相高效液相质谱(LC/MS)的方法对纯化的水稻线粒体蛋白进行分离,再用胰蛋白酶消化,然后进行串联质谱分析(MS/MS)。查找水稻基因组的开放阅读框架译码和6个表达序列标签(EST,Expressing sequence tags)译码,与质谱分析所得数据进行比对,鉴定了其中149个蛋白点(91个非冗余基因的产物),包括亲水/疏水蛋白、强酸/碱性蛋白和大分子蛋白(分子量为6.7~2.52kD)的序列。确定了85个蛋白的功能,包括线粒体的许多主要的功能蛋白。Millar等(2001年)分析拟南芥组培细胞线粒体蛋白双向凝胶电泳的结果,有大约100个高丰度蛋白和250个低丰度蛋白。用MALDI-TOF-MS分析其中170个蛋白点。查找数据库中拟南芥基因组的编码序列,鉴定了其中的91个蛋白。又通过序列比较鉴定了这91个蛋白中81个蛋白的功能。这些功能包括呼吸电子传递链,三羧酸循环,氨基酸代谢,蛋白质的输入、加工与组装,转录,膜转运和抗氧化防御。Werhahn和Braun(2002年)联合应用3种不同的凝胶电泳方法鉴定了线粒体蛋白质组的部分蛋白。首先,用蓝色非变性聚丙烯酰胺凝胶电冰(B1ue-native po1yacrylamide gel electrophoresis)分离线粒体蛋白质复合体。用电洗脱法完全洗去蛋白质中的考马斯亮蓝。然后用等电聚焦,最后用十二烷基磺酸钠(SDS)聚丙烯酰胺凝胶电泳分离蛋白质复合体的亚基。该方法的可行性已通过分离ATP合酶复合体、细胞色素C还原酶复合体和线粒体外膜移位酶前体蛋白而得到验证。利用这一方法可以分离高等真核生物中蛋白亚基的异构物。