表观遗传学与组织
很久以前,当我们仍然使用限制性内切酶来识别多态性,而“表观遗传”一词被用来驳斥那些不遵循“DNA使RNA产生蛋白质”这一中心教条的古怪数据时,我们曾经认为基因是垃圾DNA海洋中碱基的线性序列。十多年前,就在人类基因组测序项目即将完成之际,“表观遗传”一词仅限于基因表达变化的稳定有丝分裂遗传的实例,而这些变化不能归因于核苷酸序列的变化。
DNA甲基化和组蛋白修饰与基因表达变化有关,大多数表观遗传学研究涉及DNA甲基化和可能的组蛋白乙酰化的分析。如今,要想成为一个持牌的表观遗传学者,就必须记住,线性核苷酸基因序列实际上是包裹在核小体上形成染色质,染色质采用开放和封闭的构象来调节基因表达。染色质本身被进一步包装成空间分离的大碱基大小的区域和亚大碱基大小的拓扑区域,这些区域很可能形成染色体的结构支架。基因调控元件,如启动子和增强子,可以通过染色质折叠(环)使它们更接近,这些环的基部被概念化为拓扑相关域(TADs)。
染色质环构象的初步研究集中在单基因座上。这些研究确定了11个锌指CCCTC结合因子CTCF在形成染色质环中的关键作用,并证明了黏着素与CTCF共同定位以稳定环。黏着蛋白还与大的启动子-增强子复合物如介体相互作用。在过去的两年里,我们对染色质组织的理解通过采用全基因组方法的研究得到了进一步的提高。已经注意到结构蛋白CTCF和/或黏着蛋白结合位点在TADS之间的边界处富集。然而,CTCF和黏合蛋白位点也存在于TADS中,因此可能不是TAD结构的唯一决定因素。
在今年6月的Cell杂志上,Phillips Cremins等人。确定了CTCF、CTCF、黏着蛋白和介体的不同组合以组合的方式工作,以细胞类型特定的方式在亚氨基丁酸酶长度标度9处功能性地组织染色质。这是使用5C-seq策略7完成的,以生成小鼠胚胎干细胞(ESCs)和神经前体细胞(NPCs)中七个基因组区域染色质相互作用的高分辨率图谱。使这项研究引人注目的是,分辨率在4kb片段水平,这大大高于以前的Hi-C技术(平均40kb分辨率),因此揭示了亚拓扑关联域。
结果表明,一些TADs在不同的细胞类型中是不变的,但在亚megabase水平的TADs中发现了染色质组织的许多细胞特异性差异。
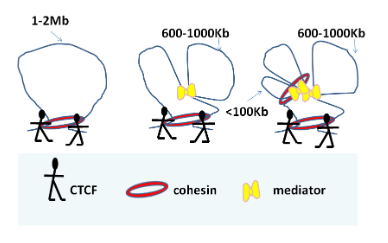
短基因组距离(<100kb)的相互作用主要发生在启动子和增强子之间,并由黏着蛋白和介体连接。在中距离(<300)CTCF处,黏着蛋白和介体常常是桥接蛋白,而在600-1000kb处,介体复合体似乎将环连接在一起。单用CTCF或CTCF与黏着素桥联的相互作用大于1MB。观察ESC和NCPs之间所观察到的染色质拓扑层次差异的范例是否适用于所有细胞类型将是一件有趣的事情。我的预测是会的。事实上,今年早些时候Seitan等人发现,非循环胸腺细胞中的黏着蛋白的缺失对基因组Mb区化的形成没有影响,但确实会影响特定的启动子-增强子相互作用,10这与上述在较短基因组距离内的相互作用的观察相符。有趣的是,亚甲基水平上相互作用的减少增加或加强了较大区域内的相互作用,这表明一个区域的构象变化影响了更宽区域的构象。
染色质地形图异常在疾病中的发生率尚不清楚。由于染色质组织是所有细胞的普遍特征,因此影响染色质组织的基因突变预计是罕见的。粘蛋白亚单位和相关蛋白参与了一系列发育障碍,统称为粘蛋白病。最著名的黏液病是Cornelia-de-Lange综合征(CdLS)。尽管在有丝分裂过程中,黏着蛋白以其将姐妹染色单体结合在一起的作用而闻名,但大多数CdLS患者在基因调控方面表现出改变,而不是有丝分裂缺陷。Bickmore实验室发现NIPBL,一种在S期将黏着蛋白复合物装载到染色质上的蛋白质,可能在调节高阶染色质结构中起作用,与CTCF和黏着蛋白无关。NIPBL基因突变的CdLS患者显示染色质失活,荧光原位杂交可见。直到最近才有CTCF突变的报道。Gregor等人现在已经在智力障碍和生长缺陷的个体中发现了CTCF基因的突变。这些突变导致CTCF的功能性单倍体缺失,来自受影响患者的转录组数据确定了几个下调基因。咨询ChIA-PET相互作用数据显示,这些患者中下调的基因可能是由于染色质拓扑结构的改变。
甚至没有提到癌症或复杂的获得性疾病,全球表观遗传基因调控被破坏,很明显,我们即将了解异常染色质构象对疾病形成的贡献。下一步是开发易于使用的检测方法,以识别各种患者组织样本上的异常染色质构象。