寄生虫引起的行为改变和不育有联系吗?
寄生虫感染可导致宿主发生多种变化。这些变化都是由同一个机制引起的,还是由几个原因引起的?一种带刺的蠕虫和它的寄主虾被用来探索这个问题。

一个复杂的生命周期涉及多个宿主是寄生动物的共同特征。这赋予了许多优势,但也带来了一个问题——如何从一个主机到另一个主机。有些寄生虫的生命周期阶段是活动的,主动寻找寄主,另一些则使用寄主(载体)来传递它们,许多寄主通过食物链相连,当它们当前的寄主被吃掉时,它们被传播到下一个寄主。这就是所谓的营养传递。
许多营养性传播的寄生虫都会耍花招,以增加宿主被吃掉的机会;但只有当它们发展到能够感染下一个宿主的阶段时。例如,当被感染时,它们的宿主可能会失去躲避捕食者的行为,甚至以吸引捕食者的方式行为。
棘头虫
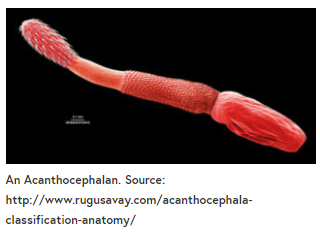
一组寄生虫——棘头虫,已知会引起无脊椎动物宿主的行为改变。成年后,棘头类动物栖息在各种脊椎动物的内脏中,它们的刺状头部长鼻通过肠壁来固定自己。
卵子进入环境,如果被脊椎动物宿主捕食的物种吃掉,棘头动物就有可能完成其生命周期。其中一个例子是多形性微小体。
多形微小体
如果一个微小疟原虫卵被一只虾吞下,它将在肠道孵化,钻入体腔,并通过棘皮阶段发展成一个桶形阶段,即囊棘。

如果受感染的虾被水禽吃掉,囊尾蚴就会在鸟的肠道中发育成一种成熟的蠕虫。
感染的影响
如果虾感觉到有捕食者的存在,它们通常会潜到池塘或溪流的底部。那些含有棘皮绦虫的昆虫也有很强的趋地性,这种行为可以保护正在发育中的寄生虫。然而,一旦寄生虫变成囊棘虫,虾的行为就会发生戏剧性的变化。它们表现出负向的地理趋同性,游到水面并依附在漂浮的植被上,从而暴露在捕食水禽的环境中(见之前的BugBitten博客)。
对这种感染的一项研究表明,低氧和与寄生虫无氧呼吸有关的排泄物可能是造成这些变化的原因。
除了表现出行为上的变化外,感染这种寄生虫的雌虾的卵不会发育,雄虾的繁殖成功率也不会降低。
基本机制
寄生虫引起的繁殖障碍和行为特征的改变在受感染的无脊椎动物中都很常见,但大多是单独研究的。这些影响背后的机制仍不清楚。特别是,我们不知道不同的表型改变是由宿主生理的单一破坏引起的,还是由多种原因引起的。
法国勃艮第大学Franche Comté的同事们通过研究小虾Gammarus pulex中的微小疟原虫感染来研究这个问题。
寻找链接
这项研究观察了与相对时间进程相关的行为变化和不育,以及它们之间的任何相关性。虾是从野外采集的。利用显微镜,通过虾的透明角质层观察寄生虫,观察其发育阶段,并确定虾的性别。随着采集季节从2月到4月的推移,随着棘刺成熟,棘刺感染的数量从63%下降到0%。
行为变化
分别对感染和未感染的虾在长玻璃柱中游泳进行监测,以确定它们的趋地性。将发育阶段相似的虾类与未感染的虾类进行比较。在所有情况下,雄性和雌性的行为方式相同,未受感染的虾类的趋地性得分随时间变化没有差异。
感染虾的情况则大不相同,证实了之前几项研究的观察结果。早、棘皮期感染的虾类比未感染的虾类具有更强的趋地性。那些长有囊尾蚴的动物有负的趋地性;这种行为会使它们接近野禽,如野鸭。这种行为的转变发生了好几天。
生殖潜能

雄虾用特殊的爪子从后面抱住产卵的雌虾,并在交配前携带它们几天。交配行为的评估方法是将一只感染的雄性和一只接受交配的未感染的雌性放在一起,让它在一个小时内抓住她。受感染的雄性在交配行为上表现出非常显著的下降。
通过检查这些爪子的大小和计算精子储备来评估男性的生殖特征。虽然棘爪大小不受囊棘的影响,但精子储备在感染阶段显著增加,但不会提前。如果没有女性授精,精子可能正在积聚。
通过计算育雏袋中的卵子或胚胎数量来评估雌性生殖能力。感染并没有造成卵子数量的差异,然而,感染的雌性,无论感染的阶段,几乎没有卵子发育。作者用阉割这个词来描述这一点。尽管完全不正确,因为这里没有涉及到对生殖组织的不可逆转的破坏,但寄生阉割一词通常用于描述对宿主生殖有负面影响的感染(见我在寄生虫学进展中的讨论,29)
相关性
行为和阉割的时间变化没有差异,行为变化的强度和阉割的强度之间也没有相关性,这并不是导致这些现象的单一机制原因。然而,作者指出,寄生虫或宿主免疫反应所分泌的产物可能会随着时间的推移而累积,并且这些性状对不同浓度的这些产物可能有不同的反应。出租车的行为甚至可能对不同的浓度有双相反应。他们还没有排除解释不同表型改变之间联系的共同机制。
显然需要对这一模型和其他模型进行进一步的研究,因为确定这些机制对有关寄生虫操纵宿主的整个辩论具有重要意义。