我被这篇关于 m6A 修饰的文章折服了……
康测科技
最近小编读到一篇关于 m6A 修饰的文章,完全被震撼和折服了!!思路之清晰、手段之全面、机制之详尽,让小编甘愿奉上自己的膝盖……毫不夸张的说够得上 Cell 级别(但是为什么没发 Cell 呢?),其他生物学过程/领域完全或部分套用这篇文章的思路,10 +文章妥妥的!!强烈建议收藏!!长文预警!!

研究背景和待解决的科学问题
癌细胞的转移是癌症致死的最主要原因,超过 90% 的癌症死亡与转移有关。EMT(上皮细胞-间质细胞转化)则是癌细胞转移过程中极其重要的一个步骤:上皮细胞在变成间充质干细胞的过程中,丧失极性并获得入侵能力。
EMT 过程的一个重要的标志是 E-cadherin 表达的丧失。EMT 是一个被研究的的非常深入的生物学过程,前期的研究表明转录因子 Snail 通过招募表观遗传修饰蛋白如 LSD1(赖氨酸去甲基化酶)和 Suv39 H1(组蛋白甲基化转移酶)调控 CDH1 基因的瞬时和长期表达水平。但是在 RNA 表观遗传调控是否在 EMT 中发挥作用,到目前还没有报道。
因此,本文要解决的科学问题,就是回答 RNA 表观遗传是否参与了 EMT 过程;如果有,那么是否存在核心的调控基因,是谁?
RNA 表观遗传目前研究的最多的就是 m6A 的修饰了。M6A 修饰前期已经被报道广泛参与各种生物学过程,如热休克反映、紫外诱导的 DNA 损伤、神经元功能与性别决定等等,在乳腺癌、肝癌等肿瘤的发生发展中的作用也有报道。因此作者选择对 EMT 过程中的 m6A 修饰的功能和作用机制进行研究。有三大类蛋白参与 m6A 的修饰:
第一类是负责修饰的甲基化转移酶,如 METTL3 / 4、RBM15 等;
第二类是去甲基化酶,如 FTO、ALKBH5;
第三类是效应蛋白,包括 YTHDFs、IGF2BPs 等。
这三类蛋白是研究 m6A 功能的重要的工具。
研究思路解析
EMT 过程中是否有 m6A 的参与?
EMT 过程中 m6A 调控的核心基因的鉴定;
m6A 对目的基因的调控机制研究;
m6A 对目的基因的调控是否与表型相关?
1.EMT 过程中是否有 m6A 的参与?
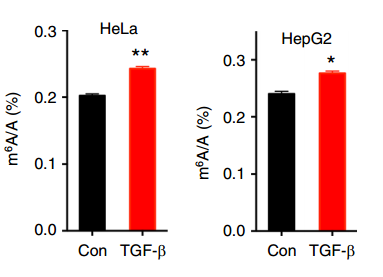
为了回答这个问题,作者使用 TGF-b 诱导包括 Hela、HepG2 等多种细胞的 EMT,并使用 LC-MS/MS 对诱导前后 RNA 的 m6A 水平进行了鉴定。结果表明诱导之后,细胞内的 m6A 水平有了显著提高,说明 EMT 过程中 m6A 的修饰水平发僧了显著变化。
那么 m6A 的这种变化是否是 EMT 过程中必要的呢?作者对甲基化转移酶 METTL3 进行部分敲除 (Mettl3Mut/−),降低 m6A 的修饰水平,然后使用 TGF-b 进行诱导,观察 m6A 修饰水平降低对 EMT 过程是否有影响。结果表明:
1.Mettl3Mut/−细胞的损伤修复和侵袭能力都有了显著下降;
2.Mettl3Mut/−细胞中 mesenchymal 标志物如 MMP2 和 FN 的 RNA 和蛋白表达水平都出现显著下降、而 epithelial 标志物 E-Cad 的 RNA 和蛋白水平则显著升高。
这些结果表明 METTL3 缺失,抑制了细胞的 EMT。
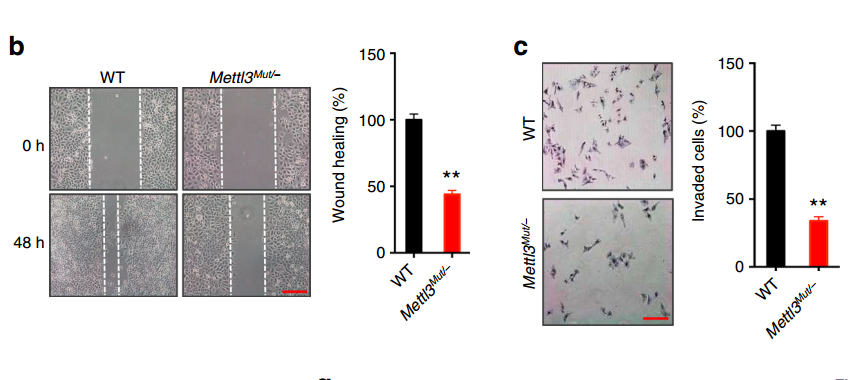
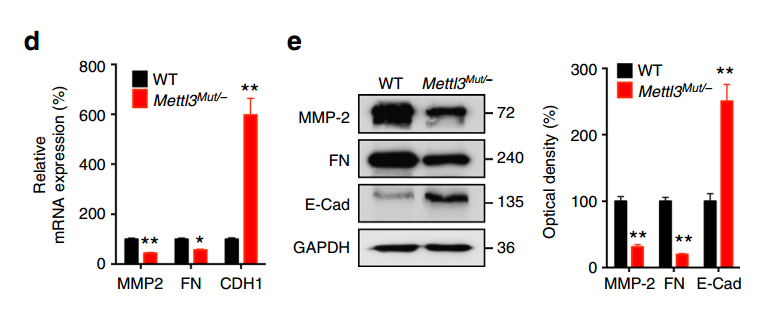
上述 METTL3 缺失对 EMT 影响的结果非常显著,但是作者尚不能确定是 m6A 水平降低引起的:因为 METTL3 虽然是一个甲基化转移酶,但它还有其他的生物学功能,因此无法将这些作用等同于 m6A 的作用。为了进一步确认是 m6A 的贡献,作者过表达了一个去甲基化酶 ALKBH5,并对上述的标志物进行了检测,取得了和 METTL3 敲除同样的效果:
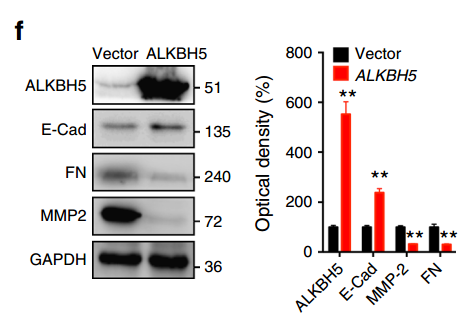
有了这个结果,已经可以下结论:m6A 参与了 EMT 的过程,并且非常重要!!
2.EMT 过程中 m6A 调控的核心基因的鉴定;
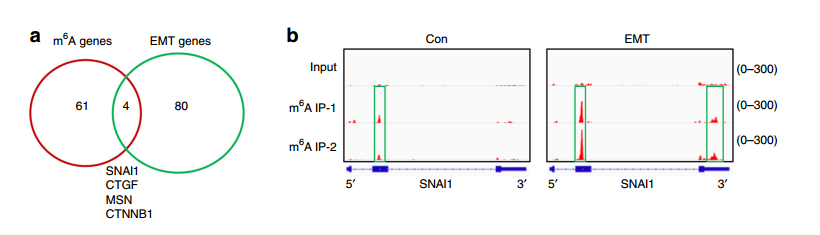
既然确认 m6A 在 EMT 中存在调控功能,因此下一步就是对被调控的基因进行鉴定了。作者使用 meRIP-seq 实验,对 EMT 前后细胞中 RNA 的 m6A 修饰状况进行了研究。发现 EMT 前后 m6A 的 motif、RNA 功能区的分布和基因组上的分布都没有显著区别,但鉴定出了 128 个 m6A 修饰水平存在显著差异(1.5Fold)的基因,这些基因的功能进行分析发现与癌细胞中的粘附连接和肌动蛋白细胞骨架的形成等 EMT 相关通路。
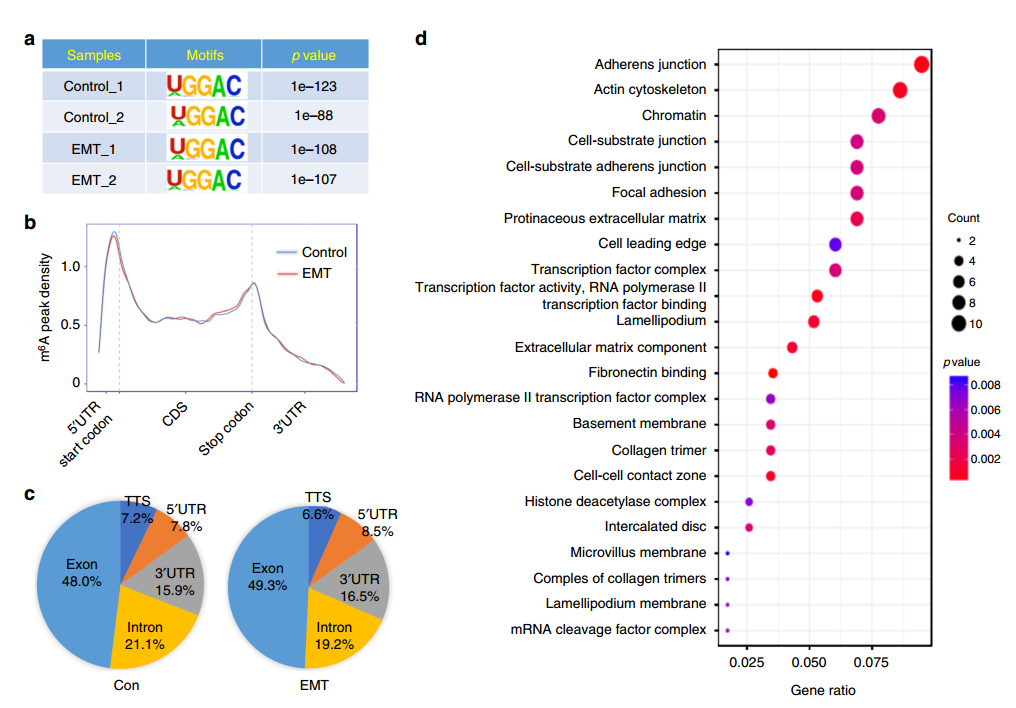
为了进一步缩小范围,找到主效应基因,作者将 meRIP-seq 中鉴定出来的 EMT 前后表达显著差异的 84 个基因和 m6A 修饰差异超过 2 倍的 65 个基因进行 overlap 分析,发现有 4 个基因 EMT 前后 m6A 水平发生了显著变化,同时表达水平也发生了显著变化,其中包括 Snail 基因。Snail 是一个已知调控 EMT 的转录因子,因此作者接下来将研究的重点放在 Snail 上。
作者首先根据 meRIP-seq 中鉴定出的 Snail 基因的 m6A 修饰位点,设计引物使用 meRIP-qPCR 进行了验证。进而使用 Western Blot 验证了 Snail 的表达水平,发现 Snail 同时受到 METTL3 和 ALKBH5 的调控,也就是 Snail 的表达水平的确受 m6A 的修饰水平调控。
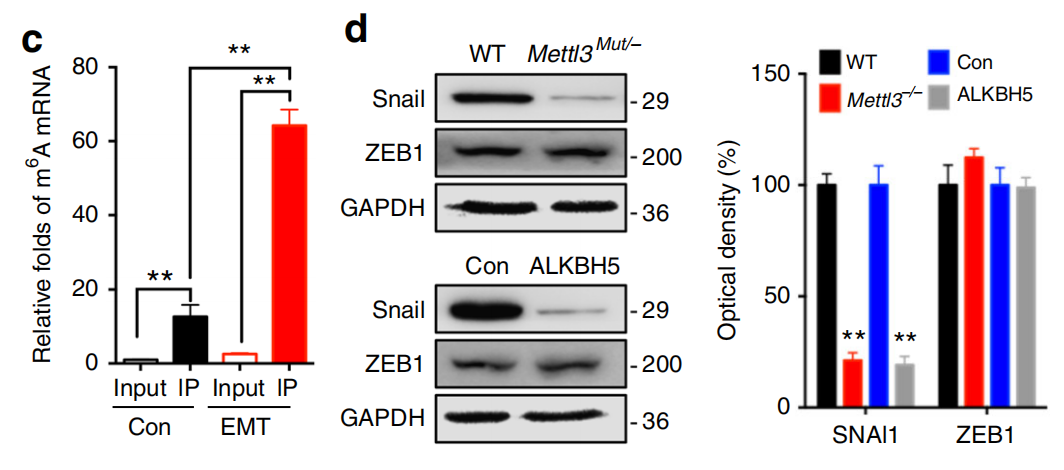
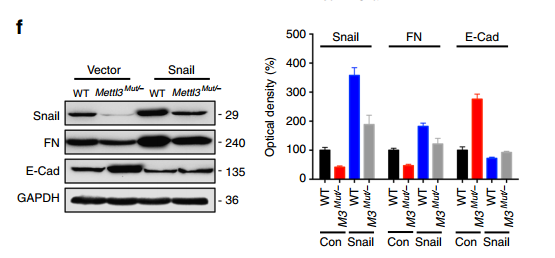
进而作者对 Snail 在 EMT 中的功能进行了验证,发现不管是从损伤修复能力还是 EMT 的蛋白标志物方面,Snail 都可以促进 EMT 的发生,重要的是 Snail 过表达可以回补 METTL3 缺失引起的 EMT deficiency。

3.M6A 对目的基因的调控机制研究;
上面的结果已经证明 Snail 受 m6A 修饰调控,那么这个调控发生在什么水平呢?由于这一部分结果太多,小编整理如下:
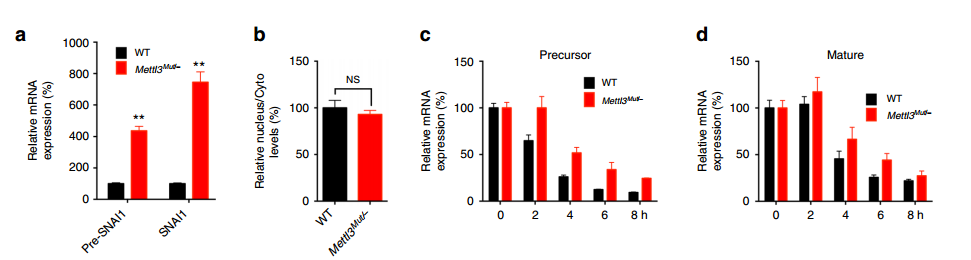
METTL3 敲除大幅上调 Snail 的 RNA 水平:
qPCR 实验验证 pre-mRNA 和 mature mRNA 的表达,发现 METTL3 敲除之后,两者的表达水平都有大幅上升
Leuciferase 实验表明 METTL3 敲除前后,Snail 的转录水平没有发生变化;
核质分离鉴定 Snail RNA 的分布,发现 METTL3 敲除前后,核质分布没有变化;
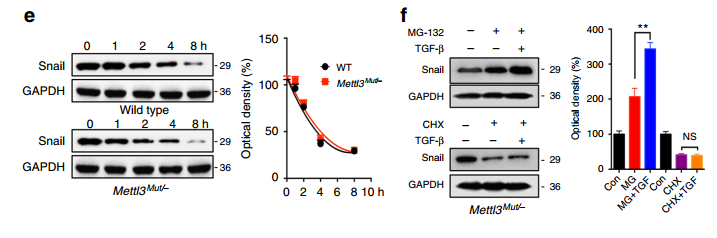
Half Life 实验表明,METTL3 敲除后,pre-mRNA 和 mature mRNA 的 stability 有了显著提高
这与蛋白水平的结果完全相反,说明 m6A 对 Snail 在蛋白水平存在更强的正调控。因此作者继续进行研究:
蛋白稳定性实验结果表明,METTL3 敲除前后对 Snail 蛋白的稳定性没有影响;
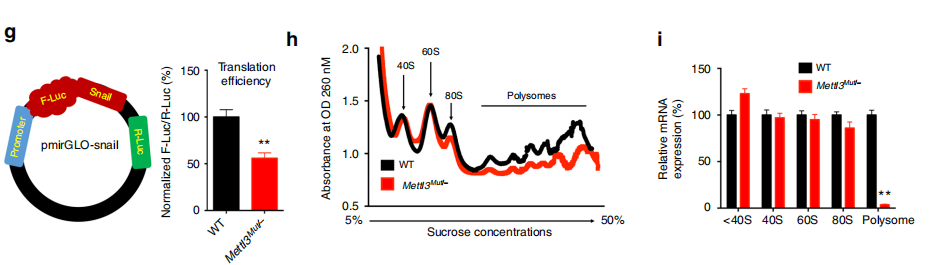
Leuciferase 实验结果表明,METTL3 敲除之后蛋白的翻译效率大幅降低;
Ribosome Profiling 实验结果表明,METTL3 敲除之后,polysome 的形成大幅降低,说明调控了翻译的延伸过程;
上述结果表明:METTL3(m6A 修饰)一方面使 Snail RNA 变得不稳定,另一方面又大幅提高翻译延伸的效率,叠加后对 Snail 蛋白水平起正调控作用。那么这个调控又是如何发生的呢?
因此作者接下来做了 2 方面的工作:
对 Snail RNA 上功能修饰位点进行确定;
鉴定介导翻译延伸效率调控的作用因子。
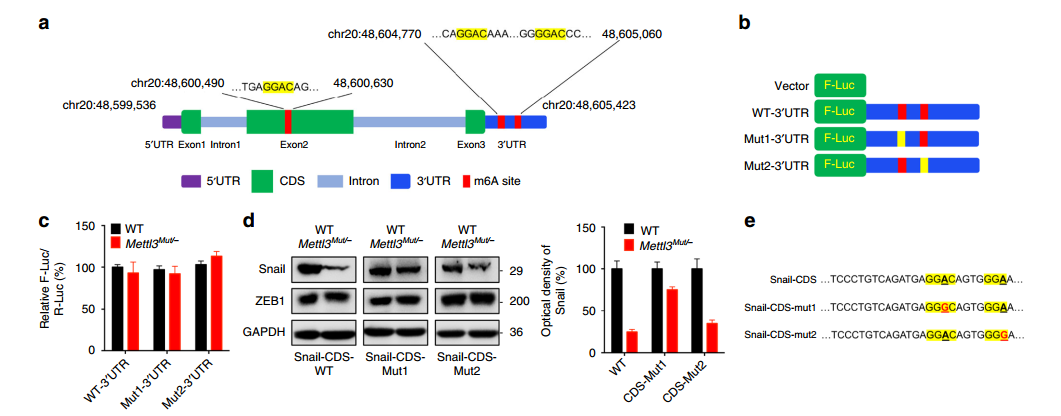
作者对 Snail RNA 上鉴定出来的 3 个 m6A 修饰位点进行了突变并检测其蛋白产物,发现:
3' UTR 的 2 个修饰位点对翻译没有影响;
- CDS 上的修饰位点对翻译效率很重要;
从而鉴定出了功能修饰位点,那么又是什么因子介导了修饰的 RNA 和翻译机器之间的互作呢?作者选择了已知的 m6A 效应蛋白(reader),YTHDF1 进行了验证:
RIP-qPCR 结果表明,YTHDF1 可以强力结合 Snail RNA,并且 METLL3 敲除之后,结合能力显著减弱;
Western Blot 结果表明,YTHDF1 敲除之后 Snail 的蛋白水平有显著下调;
RIP-qPCR 结果表明,翻译延伸因子 eEF- 2 对 Snail RNA 有强力结合,且 METTL3 敲除之后,结合显著减弱;
Co-IP 结果表明,YTHDF1 与 eEF- 2 存在互作,且 METTL3 敲除之后,两者之间结合能力下降。
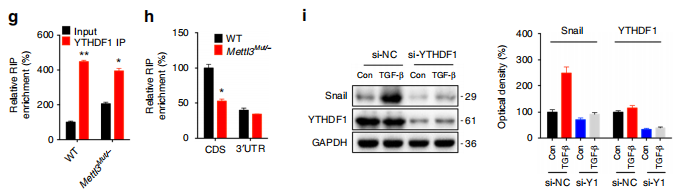
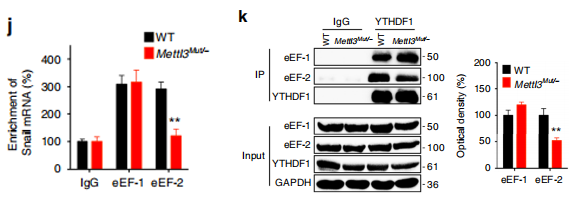
上述结果表明:m6A 对 Snail 蛋白延伸效率的调控,依赖于 YTHDF1 对其 CDS 区 m6A 位点的识别,以及 YTHDF1 与 eEF- 2 的互作。
4.M6A 对目的基因的调控是否与表型相关?
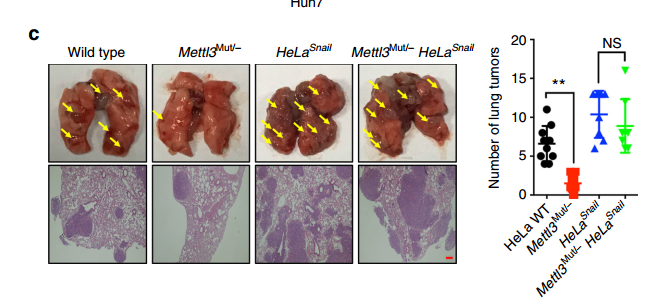
最后作者对 METTL3 -Snail 通路进行了功能验证。作者将对照或 sh-METTL3 的 Huh7 细胞注射裸鼠,成瘤后对瘤组织进行 IHC 检测,发现 METTL3 敲低后,瘤组织中 Snail,FN 和 Vim 表达降低。为体内验证 m6A 修饰对细胞转移的影响,作者将 HeLa WT,Mettl3Mut/- HeLa,HeLaSnail 和 Mettl3Mut/- HeLaSnail 稳定细胞系注射到裸鼠中。结果表明 METTL3 敲低细胞的肺肿瘤数量显着减少;而 Snail 的过表达可以回补 METTL3 敲低的抑制作用,这表明 Snail 参与了 METTL3 的体内转移。

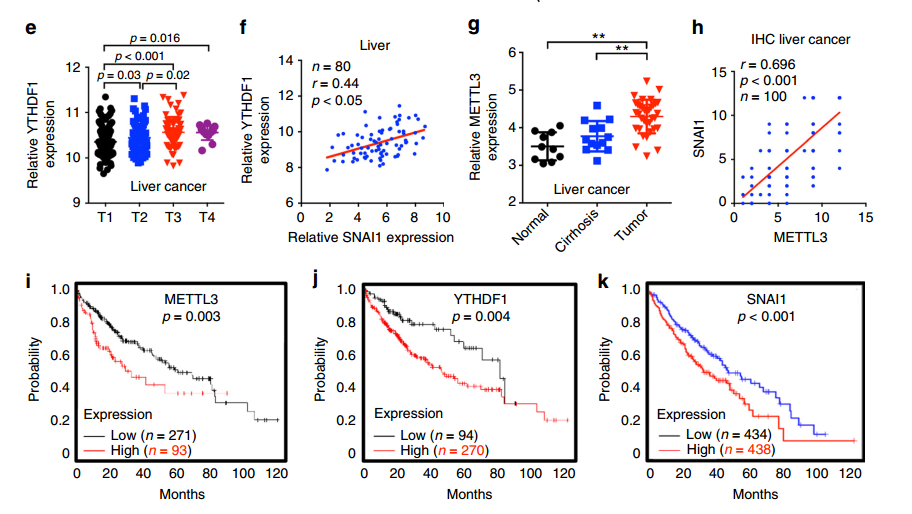
同时作者对 TCGA 数据库中肝癌的数据进行了分析,发现肝癌组织中 METTL3 和 YTHDF1 表达增加、T1 至 T4 期 YTHDF1 表达水平显着升高、YTHDF1 的表达与 Snail mRNA 的表达正相关、METTL3 从正常肝脏到肝硬化到肝癌组织的表达水平上升。另外发现 METTL3、YTHDF1 和 Snail 表达增加的肝癌患者显示总体存活率(OS)降低。
总结
先总结一下这篇文章的思路:
1. LC-MS/MS 确定 EMT 过程中有 m6A 修饰的变化;
2. 对 METTL3 和 ALKBH5 两个层面证明 m6A 在 EMT 中有功能;
3. meRIP 鉴定主要被调控的基因,聚焦到 Snail 上;
4. 对 Snail 及其 m6A 修饰对 EMT 的作用进行验证;
5. 对 m6A 修饰如何调控 Snail 基因的表达机制进行研究
- Leuciferase 确定转录活性;
- qPCR 确定 RNA 加工;
- 核质分离确定 RNA 定位;
- actinomycin D 处理,Half Life 确定 RNA 稳定性
- CHX 处理,Half Life 确定蛋白稳定性
- Ribosome Profiling 确定翻译延伸变化
- 位点突变确定功能修饰位点
- RIP-qPCR 确定 YTHDF1 对 RNA 的结合
Co-IP 确定 eEF- 2 和 YTHDF1 的互作
6. 对 METTL3 -Snail 通路的功能进行动物实验验证和大样本验证
本文的前五部分从大到小逐级深入,是研究 m6A 修饰的教科书级别思路。尤其是第五部分,覆盖了基因表达从转录到翻译全过程的研究方法,值得珍藏并借鉴。但是这篇文章为什么「只能」发 NC 呢?(以下是小编的一家之言,胡言乱语如有冒犯请装作没看见)小编认为主要是因为筛选出来的基因是 Snail:Snail 是一个已知的 EMT-inducing transcription factor,其在 EMT 过程中的调控机制已经被研究的非常深入了。

因此本文在 m6A 如何调控 Snail 表达这部分做的非常细致,但从 snail 到 EMT 就无法继续了,读起来有点头重脚轻的感觉。如果换成一个其他的不是那么 well characterized gene,Cell 妥妥的。当然,发现什么不是作者自己能决定的,这也不影响这项工作的优秀。小编仍然认为这是一篇参考价值极其高的文献!!