




你知道一个数据库的文献量有多大么? 仅仅是万方中关于期刊的文献就有 36 000 000+ 篇,而 Pubmed 中则有 27 000 000+ 篇。每一个数据库的文献量都可以用「浩如烟海」形容。当然,你根据自己需要的主题进行检索所得的结果并没有这么多。即使如此,检索的文献量少则数十上百篇,多则上千篇,难不成你还要将所有的文献都浏览一遍,才知道哪些文章对你的研究是有帮助?即使是简单的浏览标题,也能让你两眼昏花,更何况进一步阅读摘要,甚至是全文。那么,接下来的关 ...
参加了多少次海内外大大小小的学术会议,没个展示自我科研成果的壁报可怎么行?那么,怎样制作出一份精致出彩的壁报呢,今天就带大家共同学习一番。1 明确会议要求查看会议方对壁报的要求,包括内容题材、壁报的尺寸、具体的格式要求等。2 简练撰写报告出众的观点,良好的表述,与出彩的展示均是不可或缺的。同样是对研究内容的简明扼要展示,如果还不会写结构式摘要,建议你先掌握好摘要的写法。然后,过渡性地转化为壁报内容,做到简明清晰易懂,有结构 ...

对于刚入门的科研小白,免疫组化步骤较为繁琐,而且一步错,步步错,有时候明明觉得自己都按照老师们提供的条件做,可惜就是没有得到预期结果,现我将自己的些许体会分享如下。关于免疫组化的定义,原理,分类,优势等不再一一介绍(丁香园等主流平台都有介绍),我只叙述我认为「比较小」的问题,但这些往往决定了成败,遗憾的是我检索了比较多的资料,都没有详细的提及这些问题。1 取材与制片① 一定要按照标本的标本留取要求留取,譬如某组织要求固定前要 ...
写在前面的话本文是一篇脊柱领域系统评价,成文后自我评估不甚看好,理由有三:阴性结果一般不受待见中文文献偏多纳入文献质量偏低先后投稿以下七个期刊,最终被 AOTT 接受。European Spine Journal(2.132,投稿时 IF,下同)Clinnical Orthppaedics and Related Research(3.127)International Orthopaedics(2.387)Acta Orthopaedica Belgica(0 ...
我做 western 的时候,发现目前的资料都很老,网上的资料也大多互相传抄,说法各异,于是综合了目前网上所能找到的资料和个人实际经验,编辑了这份 western blot 操作步骤,内容包括主要操作步骤和所需要注意的有用的细节,并附上参考文献。我按下文的步骤操作已经做出来了稳定可重复的、背景干净的 western 条带了,屡试不爽,所以相信你也行。背景蛋白质印迹的发明者是斯坦福大学 George Stark。Neal Burnette 于 1981 年所著的 A ...
免疫共沉淀(Co-Immunoprecipitation)是以抗体和抗原之间的专一性作用为基础的用于研究蛋白质相互作用的经典方法。它可以:测定两种目标蛋白质是否在体内结合;确定一种特定蛋白质的新的作用搭档;分离得到天然状态的相互作用蛋白复合物。01 实验原理当细胞在非变性条件下被裂解时,完整细胞内存在的许多蛋白质-蛋白质间的相互作用被保留了下来。如果用蛋白质 X 的抗体免疫沉淀 X,那么与 X 在体内结合的蛋白质 Y 也能沉淀下来。目前多用 ...
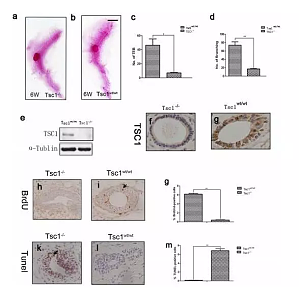
夏天到了,太平公主们该发愁了,我是多垫几层海绵呢,还是多吃木瓜呢,如此等等,各种丰胸办法都用上。其实要我来说,真没啥用,别浪费钱了,天生的,基因决定的。不行我们来看看 Nature 上的一篇文章《Tsc1 deficiency impairs mammarydevelopment in mice by suppression of AKT, nuclear ERα》,从标题就可以看出 Tsc1 缺失会影响小鼠发育,到底是怎么影响的呢?Tsc1 缺失下调了 AKT, nuclear ERα 的水 ...

网民会通过各种方式存储各种格式的影片视频,现如今,生物学家也不例外。研究人员通过使用微生物免疫系统 CRISPR-Cas,已成功将电影片段编码存储到大肠杆菌的基因组中。该技术成果于 2017 年 7 月 12 日 在《Nature》率先发布。美国马萨诸塞州哈佛医学院的合成生物学家 Seth Shipman 提到,将编码数据存储进细菌 DNA,有助于存储记录一系列事件的发生发展。Shipman 在过往研究脑神经发育时,因缺乏记录捕捉脑神经细胞运作的技术而受阻。当然 ...
实验步骤01 探针的标记如下设置探针标记的反应体系:体系为 10 微升待标记探针 (1.75 pmol / 微升) :2 微升。T4 Polynucleotide Kinase Buffer (10×) :1 微升。Nuclease-Free Water :5 微升。 ATP(3 000 Ci/mmol at 10 mCi/ml) :1 微升。T4 Polynucleotide Kinase (5-10 u / 微升) :1 微升。依次加入各种试剂,加入同位素后混匀,再加入 T4 Polynucleotide Kinase,混匀。使用水浴或 PCR 仪,37 ℃ 反应 10 分钟。加入 1 微升 ...
今年的基金评审即将进入会审阶段,相信很多人都在等待最后宣判,有些没有上会的朋友可能已经提前知道了结果。今年和 2016 年一样,我参与了 15 项青年基金的评审,这是一项很费时、费精力的工作,今年花了我 1 个星期左右的时间,审完后有一些感想,与大家分享。心急的或者不想看文字的,可以直接拉到文末看视频教程,手把手教你《基金标书的申报与写作技巧》。不管是审标书还是论文,我都是非常认真地对待的,今年和去年相比,标书质量明显提高,这个可能和各地 ...
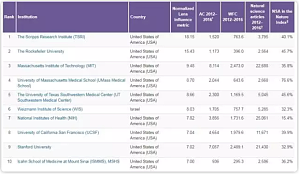
最新一期(8 月 10 日)Nature 杂志根据 Nature 指数,高质量科学指标,以及 Lens 平台的数据,排列了全球具有创新力的 200 家研究机构和高校。调查了科研论文在第三方所持有的专利中的引用情况,为理解学术研究对创新的影响这一问题带来了新思路。评判标准除了 Lens 指数,还有论文的数量和质量,以及研究机构在 InCites 中的自然科学文章。根据 Lens 开发的指标,前十位排名显示如下,其中以色列的魏茨曼科学研究所(Weizmann Institute of Sc ...
韩春雨撤稿事件近期闹到沸沸扬扬,这件事其实还告诉我们一个这样的道理:在生物科研领域,不会出现什么屌丝逆袭的情况……从到只是短短一年的时间......但我们可以注意一下在撤稿时,Nature 官方的措辞:也就是说,其实目前根本没有发现韩春雨造假的迹象,现在就盖棺定论还为时过早。韩春雨主动撤稿的最主要原因,想必并不仅仅因为实验难以被重复的科学问题,而是由于同行口诛笔伐的压力。这是韩春雨发表这篇文章的同期文章,只要看一眼作者数 ...
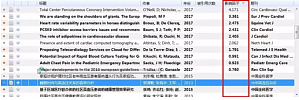
在《如何查找核心文献(一):知网高级检索》一文中,我们提到了如何在知网中查找核心文献,同样的,在万方中也能使用「被引」对检索的文献进行排序,这里就不详细介绍了(个人觉得万方没有知网做的好,万方没有计量可视化分析 —— 具体内容可以在《如何查找核心文献》中阅读)。接下来,我们就要进一步讨论:如何应用文献管理软件来筛选核心文献。一般而言,文献管理软件主要是应用于题录(包含全文)的整理,在我们写论文时方便自动插入参考文献,而不需要手动编 ...

大数据的时代,文章的受试者也开始成指数增长,动不动就是成千上万的样本。在遗传学研究中,genomAD 目前已经有超过一万五千个体的的全基因组测序数据和超过十二万人的全外显子测序数据。国际合作的大项目,如 GTEx 也有近千人死亡后捐献尸检组织的表达数据,包含了不同的组织。这对于想要做点有创意的临床研究的你来说是不是压力山大,这要积累多少年才能收那么多病人……今天给大家介绍另外一个出路,做少而精的长期随访研究,只要想法独到, ...
你是不是会有这样的经历:距离文献汇报只剩一周的时间了,但是实验累身,文献还没有开始看,一瞥到那密密麻麻的英文就心急如焚?又或者老板给你下达任务,这学期结束前必须完成一篇综述的初稿,要求阅读 80 篇以上的英文文献,英文不好的你正抓耳挠腮?别着急,接下来笔者介绍几个实用的方法,让你阅读英文文献的速度咻咻咻地提高。✎ CNKI 翻译助手CNKI 翻译助手,是「中国知网」开发制作的大型在线辅助翻译系统。用户不仅可以输入词汇还可以输入句子进 ...
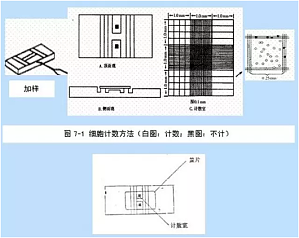
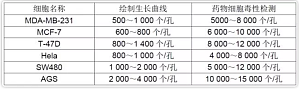
继上次我们推出初学者实验教程系列之后,很多站友发来私信,希望我们分享更多实验有关的内容。请大家继续关注我们的献给初学者栏目,会有更多大家常用的实验教程和大家见面。今天我们就来聊一聊细胞计数实验。实验用品0.4% 台盼蓝溶液、无水乙醇或 95% 乙醇溶液、脱脂棉、普通显微镜、试管、吸管、毛细吸管、细胞计数板、绸布。实验原理在细胞培养工作中,常需要了解细胞生活状态和鉴别细胞死活,确定细胞接种浓度和数量以及了解细胞存活率和增殖度,如用 ...
做了 5 年的 MTT 实验 ,耗费了上万块 96 孔板。有成功的喜悦,有失败的沮丧,有好多话要对各位实验同仁说。在这之前,我们先回顾一下 MTT 的原理及操作步骤。实验原理MTT 是一种接受氢离子的染料,可作用于活细胞线粒体中的呼吸链,在琥珀酸脱氢酶和细胞色素 C 的作用下,生成蓝色的 formazan 结晶。formazan 结晶的生成量仅与活细胞数目成正比(死细胞中琥珀酸脱氢酶消失,不能将 MTT 还原,无法形成结晶),且该结晶溶解于 DMSO。利用酶标仪测定波长 490 nm 处的光密度 OD ...
大家好,我是红领巾。下面我要讲述的是一个没有小三的故事,一个只有两个蛋白的爱情。首先简要介绍一下 GST pull-down 技术,高大上的说法是通过已经标记的 GST 标签蛋白,从细胞裂解液中钓出与之相互作用的蛋白,用以确定已知蛋白与钓出蛋白或已纯化的相关蛋白间的相互作用关系。通俗地讲就是 GST 标签的 A 蛋白(GST-A)与另一标签的 B 蛋白(作者君用的是 His 标签纯化的体外蛋白 His-B)是否相爱的故事。从细胞裂解液中钓出与之相互作用的蛋白,作者君称 ...

自 2015 年 3 月起,Springer、Elsevier 等国际出版集团的 4 批集中撤稿中,涉及中国作者的论文有近 200 余篇,其中 Springer 旗下的《肿瘤生物学》就撤稿超过 100 篇,撤销原因大多是论文在同行评审报告中造假。大部分作者委托第三方中介机构代为投稿,而恰恰是这些第三方机构通过冒用本领域专家的名字申请邮件地址,在专家不知情的情况下,用虚假邮箱向期刊提供正面夸赞论文的审稿意见。图 1 人民名义中关于高校科研领域存在的潜规则的一段对话买版面 ...

说到中国风你会想到什么?青花瓷、水墨画、大熊猫、万里长城、太极 Kongfu、龙凤、鸟巢、哪吒、孙悟空...... 近些年来,它们陆续出现在各大顶级学术期刊的封面上,让世人新奇,让国人自豪!今天就来看下有哪些中国神兽已上线。01 青花瓷镶龙青花瓷镶龙,龙绕神舟,《Nature》聚焦中国科学专题,论中国科学的腾飞进步与发展。主页链接http://www.nature.com/news/science-in-china-1.2009402 双龙戏火珠喜 ...

