Recombinant DNA Engineering, Or Cloning Genes In Plasmids
互联网
971
Plasmids
A DNA molecule can be amplified using the in vitro technique of PCR to obtain large number of identical molecules. Such pure DNA is needed, for example, to sequence it, to use as a probe in Southern or northern analysis, and so on. If the DNA molecule has promoter elements (to allow RNA polymerases to transcribe the DNA), one can also obtain usable amounts of pure RNA, and RNA with a coding region can be translated in vitro or in vivo to express a protein of interest.
The PCR technique however is error prone and is not practical as compared to the in vivo, cloning technique to produce large amounts of identical DNA. The DNA of interest is made to propagate in bacteria as plasmid DNA. Bacteria have the machinery to replicate the plasmid DNA with very high fidelity. Plasmid DNA is physically independent of genomic DNA and this allows for their easy isolation from bacterial cultures.

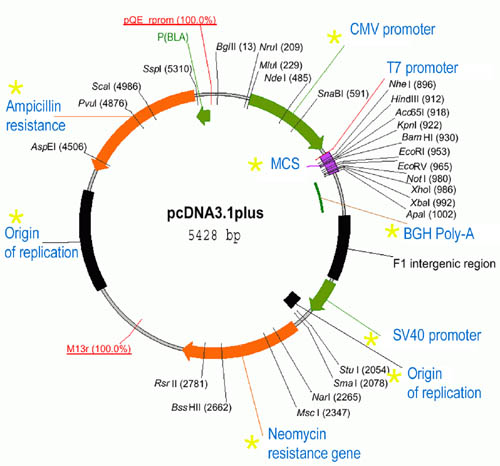
the pcDNA3.1+ plasmid
Vector plasmids used for cloning are short, circular, double-stranded DNA. They have at least one origin of replication (a DNA segment; for propagation as bacteria divide). They also have a segment encoding for resistance against some antibiotic like ampicillin. These resistance genes allow selection of bacteria carrying the plasmid as those not carrying the plasmid will not survive antibiotics in culture medium. A region of the plasmid called multiple cloning site or MCS bears sites for various restriction enzymes to allow the insertion of a DNA of interest - the insert . Plasmids usually have a promoter upstream of the MCS such that the insert gets transcribed. If the promoter is mammalian (e.g., the CMV promoter), the transcription will take place only in mammalian systems (like mammalian cell culture), but not in the bacteria. Plasmids also usually have promoter segments for phage polymerases (like T7 and SP6 RNA polymerases) such that one can use in them vitro to produce RNA. A plasmid designed for mammalian expression also usually has a transcription termination and polyadenylation signal (such as the BGH or SV40 poly-A signals) downstream of MCS. In this way the transcribed insert gets efficiently polyadenylated for stability and good translation in a mammalian cell.
A plasmid might also be designed to allow generation of fusion RNA and thus fusion proteins. For example there might be sequence for a fluorescent protein such as GFP or a peptide tag such as HA upstream of MCS (and downstream of promoter) so that an insert in frame will lead to the generation of a fusion protein, an N-terminal tagged (with GFP or HA in these examples) protein of interest.
Insertion of DNA of interest in a plasmid
Pure DNA of interest is obtained by PCR or from another plasmid and is inserted in the vector plasmid by ligation or recombination or even PCR. This however is not efficient. As a result a large number of empty plasmids remain in the reaction. If they are circular (e.g., in the ligation method, the plasmid is first opened up, linearized , at the cloning site using restriction enzymes; these linearize molecules can self-ligate ), the antibiotic resistance gene will allow bacteria carrying empty vector plasmids to survive the selection.
Transformation
The reaction used to create recombinant DNA molecules (ligation reaction, etc.) is used to transform bacteria. The transformation efficiency is not important if one already has a pure plasmid with the insert and just want to transform to grow more of the plasmid.
Transformed bacteria are plated on a culture plate that has antibiotics. After 15-20 hours, colonies (each colony represents one bacterium from the transformation process) are picked up. These colonies though they survive the antibiotic need not necessarily have a plasmid with the insert. One therefore screens for the right colonby or clone by various methods - PCR, restriction digestion of plasmid DNA, Southern blotting, and so on. Some cloning procedures have ingenious selection criteria. E.g., cloning of PCR products in pCR2.1 vector ( TA cloning ) allows selection of the right clone by blue-white screening . The vector has a galactosidase encoding gene which gets interrupted by the insert. Colonies that carry the plasmid but do not have an insert carry the functional galactosidas gene and thus appear blue when grown in presence of the X-gal chemical.
When a (or many) correct colonies are identified, they can be grown up in large cultures to make large amounts of plasmids ( maxipreps ). The bacteria can be frozen in 20% glycerol at -80 degrees and used for inoculating cultures in future.
The plasmids may be sent for sequencing to confirm the identity of the insert. Thorough sequencing of the insert may be needed to confirm that its sequence is as expected.
Transfection
Eukaryotic cells can be transfected with a plasmid to generate RNA for RNA interference in the cell, for synthesis of a particular protein, etc. (If the plasmid has a bacterial promoter, the protein can be made inside the bacteria itself.)
Plasmids cannot propagate in eukaryotic cells unless they are integrated (inserted in chromosmes) in the genome. As a result, and also because of degradation, transfected plasmid DNA dilutes out over 5-6 rounds of cell replication. The transient transfectants can be cultured for longer duration during which stable integration can take place. Integration is a rare phenomemon, but if there are markers for it, one can select stable cell lines.
Usually the presence of a drug resistance gene (such as one to inactivate neomycin or G418) on the plasmid allows for such selection. However, drug resistance necessarily does not mean the protein of interest is indeed expressed. One therefore has to pick many clones and confirm protein expression by western analysis, etc.
Some plasmids allow for generation of a fusion, bicistronic message. In such cases, the gene of interest is transcribed but in the absence of a termination signal, the transcription from the plasmid extends beyond the insert and another gene such as GFP that is downstream gets transcribed as well. These transcripts are bicistronic (two coding regions: gene of interest and GFP in the example). An IRES or internal ribosomal entry site upstream of the second gene (GFP) allows the translation of the second coding region (GFP) independently of translation of the first one. A clone that expresses high GFP thus suggests high expression of the first protein as well (indirectly, both proteins coming from same transcript). High expressors can thus be FACS-sorted from the drug resistant, stable population.