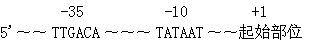启动子
互联网
启动子是位于结构基因5,端上游的一段DNA序列,能够指导全酶(holoenzyme)同模板正确结合,活化RNA聚合酶,启动基因转录。全酶是指酶蛋白及其辅酶构成的有功能的复合物。RNA,聚合酶的核心酶虽可合成RNA,但不能找到模板DNA上的转录起始位点,只有带σ因子的全酶才能专一地同启动子结合。RNA聚合酶沿着模板前进,直到终止子,转录产生一条RNA链。通常把基因转录起点前面即5’端的序列称为上游(upstream),起点后面即3’端的序列称为下游(downstream)。并把起点的位置记为十1,下游的核苷酸依次记为 2, 3,……,上游方向依次记为—1,—2,—3,……。
RNA聚合酶同启动子结合的区域称为启动子区。将各种原核基因同RNA聚合酶全酶结合后,用DNase I水解DNA,最后得到与RAN聚合酶结合而未被水解的DNA片段,这些片段有一个由5个核苷酸(TATAA)组成的共同序列,以其发现者的名字命名为Pribnow框(Pribnowbox),这个框的中央位于起点上游10bp处,所以又称—10序列(—10 sequence),后来在—35 bp处又找到另一个共同序列(TTGACA)。Hogness等在真核基因中又发现了类似Pribnow框的共同序列,即位于—25~—30 bp处的TATAAAAG,也称TATA框(TATAbox)。TATA框上游的保守序列称为上游启动子元件(upstream promoter element,UPE)或上游激活序列(uptreamactivatingsequence,UAS)。另外在—70~—78 bp处还有一段共同序列CCAAT,称为CAAT框(CAAT box)
原核生物中—10区同—35区之间核苷酸数目的变动会影响基因转录活性的高低,强启动子一般为17±1 bp,当间距小于15 bp或大于20 bp时都会降低启动子的活性。
在真核基因中,有少数基因没有TATA框。没有TATA框的真核基因启动子序列中,有的富集GC,即有GC框;有的则没有GC框。GC框位于—80~—110bp处的GCCACACCC或GGGCGGG序列。
TATA框的主要作用是使转录精确地起始;CAAT框和GC框则主要是控制 转录起始的频率,特别是CAAT框对转录起始频率的作用更大。如在TATA框同相邻的UPE之间插入核苷酸,也会影响转录使之减弱。
为什么RNA聚合酶能够仅在启动子处结合呢?显然启动子处的核苷酸顺序具有特异的形状以便与RNA聚合酶结合,就好像酶与其底物的结构相恰恰适合一样。将100个以上启动子的顺序进行了比较,发现在RNA合成开始位点的上游大约10bp和35bp处有两个共同的顺序,称为-10和-35序列。这两个序列的共同顺序如下,-35区“AATGTGTGGAAT”,-10区“TTGACATATATT”。大多数启动子均有共同顺序(consensus sequence),只有少数几个核苷酸的差别。
-10序列又称为Pribnow盒(原核生物)。在真核生物中相应的序列位于-35bp处,称为TATA盒,又称为Goldberg-Hognessbox,是RNA聚合酶Ⅱ的结合部位。-10和-35这两个部位都很重要:[1]RNA聚合酶能和-35和-10序列中的碱基和DNA主链中的磷酸基相接触;[2]离开共同顺序较远的启动子的活性亦较弱;[3]最重要的是,破坏启动子功能的突变中有75%都是改变了共同顺序中的碱基,其余25%亦为离共同顺序较近的。-35和-10序列相距约20bp,即大致是双螺旋绕两圈的长度。因为这两个结合区是在DNA分子的同一侧面,可见此酶是结合在双螺旋的一面。可以想像,它能"感觉到每个结合区的沟底中碱基所产生的特异形状。"
原核生物亦有少数启动子缺乏这两个序列(-35和-10)之一。在这种情况下,RNA聚合酶往往不能单独识别这种启动子,而需要有辅助蛋白质的帮助。可能是这些蛋白质因子与邻近序列的反应可以弥补启动子的这个缺陷。
在真核生物中,在转录起始位点上游70-80bp处有CAAT顺序,也称为CAAT盒。这一顺序也是比较保守的共同顺序:GCCTCAATCT。RNA聚合酶Ⅱ可以识别一顺序。近年来在对家兔β珠蛋白基因CAAT顺序的研究中发现,用人工方法诱导CAAT顺序发生突变使家兔β珠蛋白基因的转录水平降低。
启动子中的-10和-35序列是RNA聚合酶所结合和作用必需的顺序。但是附近其他DNA顺序也能影响启动子的功能。例如,在核糖体RNA合成的起始位点的上游50到150核苷酸之间的顺序就是对启动子的完全活性所必需的。如果这一段DNA顺序缺失并由其他外来DNA所取代(例如克隆在质粒DNA中的rRNA基因),则转录起始的频率将降低10倍。同样,在其他情况下,远隔部位的富有AT的DNA顺序被认为能增进转录起始的频率。有时候上游顺序可以是某些能直接激活RNA聚合酶的"激活蛋白"的结合部位。但是,上游顺序往往有另外的功能。例如上游顺序可以吸引拓扑异构酶,后者可导致结合的局部产生有利于转录起始的超螺旋状态。上游顺序所引起的DNA结构的微细变化可能在双螺旋上被传导到相当远的距离,因此上游顺序的变化可以影响到-10和-35区的DNA结构细节。

![Cas9 SmartNuclease AAVS 1-gRNA靶向载体[EF 1a启动子]-Cas9 SmartNuclease AAVS1-gRNA Targeting Vector [EF1a promoter]](https://img1.dxycdn.com/p/s14/2025/0226/793/4490333375672954981.jpg!wh200)