转座子
互联网
4083
转座子是细菌细胞里发现的一种复合型转座因子,这种转座因子带有同转座无关的一些基因,如抗药性基因;它的两端就是IS,构成了“左臂”和“右臂”。两个“臂”可以是正向重复,也可以是反向重复。这种复合型的转座因子称为转座子(trans—poson,Tn)。这些两端的重复序列可以作为Tn的一部分随同Tn转座,也可以单独作为IS而转座。Tn两端的IS有的是完全相同的,有的则有差别。当两端的IS完全相同时,每一个IS都可使转座子转座;当两端是不同的IS时,则转座子的转座取决于其中的一个IS。Tn有抗生素的抗性基因,Tn很容易从细菌染色体转座到噬菌体基因组或是接合型的质粒。因此,Tn可以很快地传播到其他细菌细胞,这是自然界中细菌产生抗药性的重要来源。
两个相邻的IS可以使处于它们中间的DNA移动,同时也可制造出新的转座子。Tn10的两端是两个取向相反的IS1O,中间有抗四环素的抗性基因(TetR),当TnlO整合在一个环状DNA分子中间时,就可以产生新的转座子。当转座子转座插人宿主DNA时,在插入处产生正向重复序列,其过程是这样的:先是在靶DNA插入处产生交错的切口,使靶DNA产生两个突出的单链末端,然后转座子同单链连接,留下的缺口补平,最后就在转座子插入处生成了宿主DNA的正向重复。已知的转座因子的转座途径有两种:复制转座和非复制转座。
1.复制转座(replicative transposition) 转座因子在转座期间先复制一份拷贝,而后拷贝转座到新的位置,在原先的位置上仍然保留原来的转座因子。复制转座有转座酶(transposase)和解离酶(resolvase)的参与。转座酶作用于原来的转座因子的末端,解离酶则作用于复制的拷贝。TnA是复制转座的例子。
2.非复制转座(non-replicative transposition) 转座因子直接从原来位置上转座插入新的位置,并留在插入位置上,这种转座只需转座酶的作用。非复制转座的结果是在原来的位置上丢失了转座因子,而在插入位置上增加了转座因子。这可造成表型的变化。
保留转座(conservative transposition)也是非复制转座的一种类型。其特点是转座因子的切离和插人类似于入噬菌体的整合作用,所用的转座酶也是属于入整合酶(integrase)家族。出现这种转座的转座因子都比较大,而且转座的往往不只是转座因子自身,而是连同宿主的一部分DNA一起转座。 非复制转座可以是直接从供体分子的转座子两端产生双链断裂,使整个转座子释放出来,然后在受体分子上产生的交错接口处插入,这是“切割与黏接”(“cut and paste")的方式。另一种方式是在转座子分子同受体分子之间形成一种交换结构(crossover structure),受体分子上产生交错的单链缺口,与酶切后产生的转座子单链游离末端连接,并在插入位点上产生正向重复序列;最 后,由此生成的交换结构经产生缺口(nick)而使转座子转座在受体分子。供体DNA分子上留下双链断裂,结果 或是供体分子被降解,或是被DNA修复系统识别而得到修复。
在复制转座过程中,转座和切离是两个独立事件。先是由转座酶分别切割转座子的供体和受体DNA分子。转座子的末端与受体DNA分子连接,并将转座子复制一份拷贝,由此生成的中间体即共整合体(cointegrat,)有转座子的两份拷贝。然后在转座子的两份拷贝间发生类似同源重组的反应,在解离酶的作用下,供体分子同受体分子分开,并且各带一份转座子拷贝。同时受体分子的靶位点序列也重复了一份拷贝。
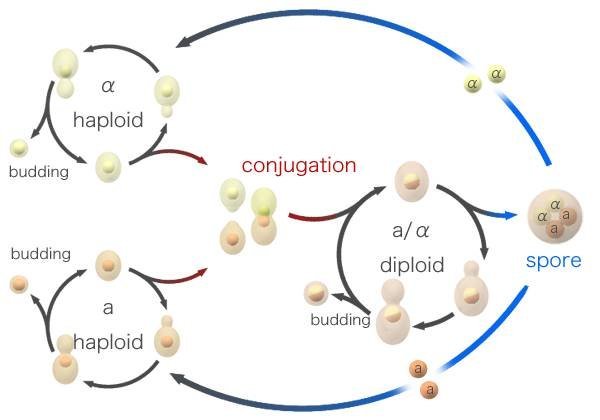
酵母接合型的相互转换也是复制转座所产生。酿酒酵母(Saccharomvcescerf—visiae)的生命周期中有双倍体细胞和单倍体细胞两种类型。单倍体细胞则有a型和α型两种接合型(mating type)。单倍体酵母是a型还是α型,由单个基因座MAT所决定。MAT有一对等位基因MAT。和MATα,在同宗接合(homothallic)的酵母菌株中,酵母菌十分频繁地转换其接合型,即从a转换成α,然后在下一代又转换为a。这种转换和回复的频率已远远高于通常的自发突变,表明这不是通常的突变机制。现在已经知道,在MAT基因座两侧有两个基因带有MATα和ATα的拷贝,这就是HMLα和HMRα基因。这两个基因贮存了两种接合型等位基因,当转座给MAT基因座时就发生了接合型的转换。因此,MAT基因座是通过转座而转换其接合型的。MAT基因座的序列转换成另一个基因的序列,这种机制称为基因转换(gene convertion)。
两个相邻的IS可以使处于它们中间的DNA移动,同时也可制造出新的转座子。Tn10的两端是两个取向相反的IS1O,中间有抗四环素的抗性基因(TetR),当TnlO整合在一个环状DNA分子中间时,就可以产生新的转座子。当转座子转座插人宿主DNA时,在插入处产生正向重复序列,其过程是这样的:先是在靶DNA插入处产生交错的切口,使靶DNA产生两个突出的单链末端,然后转座子同单链连接,留下的缺口补平,最后就在转座子插入处生成了宿主DNA的正向重复。已知的转座因子的转座途径有两种:复制转座和非复制转座。
1.复制转座(replicative transposition) 转座因子在转座期间先复制一份拷贝,而后拷贝转座到新的位置,在原先的位置上仍然保留原来的转座因子。复制转座有转座酶(transposase)和解离酶(resolvase)的参与。转座酶作用于原来的转座因子的末端,解离酶则作用于复制的拷贝。TnA是复制转座的例子。
2.非复制转座(non-replicative transposition) 转座因子直接从原来位置上转座插入新的位置,并留在插入位置上,这种转座只需转座酶的作用。非复制转座的结果是在原来的位置上丢失了转座因子,而在插入位置上增加了转座因子。这可造成表型的变化。
保留转座(conservative transposition)也是非复制转座的一种类型。其特点是转座因子的切离和插人类似于入噬菌体的整合作用,所用的转座酶也是属于入整合酶(integrase)家族。出现这种转座的转座因子都比较大,而且转座的往往不只是转座因子自身,而是连同宿主的一部分DNA一起转座。 非复制转座可以是直接从供体分子的转座子两端产生双链断裂,使整个转座子释放出来,然后在受体分子上产生的交错接口处插入,这是“切割与黏接”(“cut and paste")的方式。另一种方式是在转座子分子同受体分子之间形成一种交换结构(crossover structure),受体分子上产生交错的单链缺口,与酶切后产生的转座子单链游离末端连接,并在插入位点上产生正向重复序列;最 后,由此生成的交换结构经产生缺口(nick)而使转座子转座在受体分子。供体DNA分子上留下双链断裂,结果 或是供体分子被降解,或是被DNA修复系统识别而得到修复。
在复制转座过程中,转座和切离是两个独立事件。先是由转座酶分别切割转座子的供体和受体DNA分子。转座子的末端与受体DNA分子连接,并将转座子复制一份拷贝,由此生成的中间体即共整合体(cointegrat,)有转座子的两份拷贝。然后在转座子的两份拷贝间发生类似同源重组的反应,在解离酶的作用下,供体分子同受体分子分开,并且各带一份转座子拷贝。同时受体分子的靶位点序列也重复了一份拷贝。
酵母接合型的相互转换也是复制转座所产生。酿酒酵母(Saccharomvcescerf—visiae)的生命周期中有双倍体细胞和单倍体细胞两种类型。单倍体细胞则有a型和α型两种接合型(mating type)。单倍体酵母是a型还是α型,由单个基因座MAT所决定。MAT有一对等位基因MAT。和MATα,在同宗接合(homothallic)的酵母菌株中,酵母菌十分频繁地转换其接合型,即从a转换成α,然后在下一代又转换为a。这种转换和回复的频率已远远高于通常的自发突变,表明这不是通常的突变机制。现在已经知道,在MAT基因座两侧有两个基因带有MATα和ATα的拷贝,这就是HMLα和HMRα基因。这两个基因贮存了两种接合型等位基因,当转座给MAT基因座时就发生了接合型的转换。因此,MAT基因座是通过转座而转换其接合型的。MAT基因座的序列转换成另一个基因的序列,这种机制称为基因转换(gene convertion)。