支原体概述
互联网
支原体是目前已知一类能在无生命培养基上生长繁殖的最小的原核细胞型微生物。自然界分布广泛,种类多,分为两个属:一为支原体属(Mycoplasma),有几十个种;另一为脲原体属(Ureaplasma),仅有一种。与人类感染有关的主要是肺炎支原体和解脲脲原体。
一、生物学性状
(一)形态与结构
支原体的大小为0.2~0.3um,可通过滤菌器,常给 细胞培养 工作带来污染的麻烦。无细胞壁,不能维持固定的形态而呈现多形性。革兰氏染色不易着色,故常用Giemsa染色法将其染成淡紫色。细胞膜中胆固醇含量较多,约占36%,对保持细胞膜的完整性具有一定作用。凡能作用于胆固醇的物质(如二性霉素B、皂素等)均可引起支原体膜的破坏而使支原体死亡。
支原体 基因组 为一环状以双链DNA,分子量小(仅有大肠杆菌的五分之一),合成与代谢很有限。
肺炎支原体的一端有一种特殊的末端结构(terminal structure),能使支原体粘附于呼吸道粘膜上皮细胞表面,与致病性有关。
(二)培养特性
营养要求比一般细菌高,除基础营养物质外还需加入10~20%人或动物血清以提供支原体所需的胆固醇。最适pH7.8~8.0之间,低于7.0则死亡,但解脲脲原体最适pH6.0~6.5。
大多数兼性厌氧,有些菌株在初分离时加入5%CO2生长更好。生长缓慢,在琼脂含量较少的固体培养基上孵育2~3天出现典型的“荷包蛋样”菌落:圆形(直径10~16um),核心部分较厚,向下长入培养基,周边为一层薄的透明颗粒区。此外,支原体还能在鸡胚绒毛尿囊膜或培养细胞中生长。
繁殖方式多样,主要为二分裂繁殖,还有断裂、分枝、出芽等方式,盖因缺乏细胞壁造成分裂时二个子细胞大小均所致。同时,支原体分裂和其DNA复制不同步,可形成多核长丝体。
(三)生化反应与分型
一般能分解葡萄糖的支原体则不能利用精氨酸,能利用精氨酸的则不能分解葡萄糖,据此可将支原体分为两类(见表63-1)。解脲脲原体不能利用葡萄糖或精氨酸,但可利用尿素作能源。
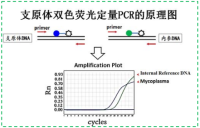
各种支原体都有特异的表面抗原结构,很少有交叉反应,具有型特异性。应用生长抑制试验(Growth inhibition test,GIT)、代谢抑制试验(Metabolic inhibition test,MIT)等可鉴定支原抗原,进行分型。
表63-1 人类支原体的主要生物学性状
|
名称 |
需要胆固醇 |
分解葡萄糖 |
分 解 |
还原四氮唑 |
吸附红细胞 |
|
|
支原体属 人型支原体 唾液支原体 发酵支原体 肺炎支原体 口腔支原体 生殖支原体 脲原体属 解脲脲原体 |
+ + + + + + + |
- - + + - + - |
+ + - - + - - |
- - - - - - + |
- - - + - ± - |
- - - + + - + |
(四)抵抗力
支原体对热的抵抗力与细菌相似。对环境渗透压敏感,渗透压的突变可致细胞破裂。对重金属盐、石炭酸、来苏尔和一些表面活性剂较细菌敏感,但对醋酸铊、结晶紫和亚锑酸盐的抵抗力比细菌大。对影响壁合成的抗生素如青霉素不敏感,但红霉素、四环素、链霉素及氯霉素等作用于支原体核蛋白体的抗生素,可抑制或影响蛋白质合成,有杀灭支原体的作用。
二、致病性与免疫性
支原体不侵入机体组织与血液,而是在呼吸道或泌尿生殖道上皮细胞粘附并定居后,通过不同机制引起细胞损伤,如获取细胞膜上的脂质与胆固醇造成膜的损伤,释放神经(外)毒素、磷酸酶及过氧化氢等。
巨噬细胞、lgG 及 lgM 对支原体均有一定的杀伤作用。呼吸道粘膜产生的SlgA抗体已证明有阻止支原体吸附的作用。在儿童中,致敏淋巴细胞可增强机体对肺炎支原体的抵抗力。
三、支原体与L型细菌的区别
L型细菌是在抗生素、溶菌酶等作用下变成的一种细胞壁缺陷型细菌,其许多特性与支原体相似,在鉴定时很有必要将二者区别开来(见表63-2)。
表63-2 支原体与L型细菌的区别
<center> <table> <tbody> <tr> <td>支原体</td> <td>L型细菌</td> </tr> <tr> <td> 自然界中广泛存在 <p class="duanluo">绝大多数生长需胆固醇</p> <p class="duanluo">在遗传上与细菌无关,且无论在什么条件下也不能变成细菌</p> <p class="duanluo">菌落较小,0.1~0.3mm</p> </td> <td> 自然界很少存在 <p class="duanluo">生长不一定需要胆固醇</p> <p class="duanluo">在遗传上与原菌相关,并可在诱导因素去除后回复为原菌</p> <p class="duanluo">菌落稍大,0.5~1.0mm</p> </td> </tr> </tbody> </table> </center>