DNA聚合酶综述
互联网
- 相关专题
- 重组DNA技术工具酶
定义
DNA聚合酶 (DNA polymerase)是细胞复制DNA的重要作用酶。
DNA聚合酶 , 以DNA为复制模板,从将DNA由5'端点开始复制到3'端的酶。
DNA聚合酶 的主要活性是催化DNA的合成(在具备模板、引物、dNTP等的情况下)及其相辅的活性。
真核细胞有5种DNA聚合酶,分别为DNA聚合酶α(定位于胞核,参与复制引发具5-3外切酶活性),β(定位于核内,参与修复,具5-3外切酶活性),γ(定位于线粒体 ,参与线粒体复制具5-3和3-5外切活性),δ(定位核,参与复制,具有3-5和5-3外切活性),ε(定位于核,参与损伤修复,具有3-5和5-3外切活性)。
原核细胞有3种DNA聚合酶 ,都与DNA链的延长有关。DNA聚合酶I是单链多肽,可催化单链或双链DNA 的延长;DNA聚合酶II则与低分子脱氧核苷酸链的延长有关;DNA聚合酶III在细胞中存在的数目不多,是促进DNA链延长的主要酶。
发现
在50年代的中期,A. Kornberg和他的同事们就想到DNA的复制必然是一种酶的催化作用,于是决心分离出这种酶并研究其结构和作用机制。为了达到这个目的,他们分离的蛋白,然后加到体外合成系统中即同位素标记的dNTP、Mg2+及模板DNA,经过大量的工作,于1956年终于发现了DNA聚合酶Ⅰ(DNA polymerase Ⅰ,DNA pol Ⅰ)原来称为Kornberg酶。以后又相续发现了DNA pol Ⅱ和DNA pol Ⅲ。开始人们以为DNA pol I是细菌中DNA复制主要的酶类,后来发现DNA pol Ⅰ的突变株照样可以复制,才清楚它并不是主角。现在已经知道在DNA复制中起主导作用的是DNA pol Ⅲ,至于pol Ⅱ的功能现在还不十分清楚。DNA聚合酶的共同特点是:
(1)需要提供合成模板;(2)不能起始新的DNA链,必须要有引物提供3'-OH;(3)合成的方向都是5'→3'(4)除聚合DNA外还有其它功能。
所有原核和真核的DNA聚合酶都具有相同的合成活性,都可以在3'-OH上加核苷酸使链延伸,其速率为1000 Nt/min。加什么核苷酸是根据和模板链上的碱基互补的原则而定的。
E.coli的DNA pol Ⅰ涉及
DNA损伤修复
,在半保留复制中起辅助的作用。DNA pol Ⅱ在修复损伤中也是有重要的作用。DNA pol Ⅲ是一种多亚基的蛋白。在DNA新链的从头合成(de novo)中起复制酶的作用。
共性
此酶最早在大肠杆菌中发现,以后陆续在其他原核生物及微生物中找到。这类酶的共同性质是:[1]以脱氧核苷酸三磷酸(dNTP)为前体催化合成DNA;[2]需要模板和引物的存在;[3]不能起始合成新的DNA链;[4]催化dNTP加到生长中的DNA链的3'-OH末端;[5]催化DNA合成的方向是5'→3'。下面首先介绍大肠杆菌的DNA聚合酶,然后简略说明一下其他原核生物的DNA聚合酶和真核生物DNA聚合酶。
功能
[1]聚合作用:在引物RNA'-OH末端,以dNTP为底物,按模板DNA上的指令由DNApolⅠ逐个将核苷酸加上去,就是DNApolⅠ的聚合作用。酶的专一性主要表现为新进入的脱氧核苷酸必须与模板DNA配对时才有催化作用。dNTP进入结合位点后,可能使酶的构象发生变化,促进3'-OH与5'-PO4结合生成磷酸二酯键。若是错误的核苷酸进入结合位点,则不能与模板配对,无法改变酶的构象而被3'-5'外切酶活性位点所识别并切除之。
[2]3'→5'外切酶活性──校对作用:这种酶活性的主要功能是从3'→5'方向识别和切除不配对的DNA生长链末端的核苷酸。当反应体系中没有反应底物dNTP时,由于没有聚合作用而出现暂时的游离现象,从而被3'→5'外切酶活性所降解。如果提高反应体系的温度可以促进这种作用,这表明温度升高使DNA生长链3'末端与模板发生分离的机会更多,因而降解作用加强。当向反应体系加入dNTP,而且只加放与模板互补的上述核苷酸才会使这种外切酶活性受到抑制,并继续进行DNA的合成。由此推论,3'→5'外切酶活性的主要功能是校对作用,当加入的核苷酸与模板不互补而游离时则被3'→5'外切酶切除,以便重新在这个位置上聚合对应的核苷酸。在某些T4噬菌体突变株中DNA复制的真实性降低,而易发生突变,从此突变株分离得到的T4DNA聚合酶的3'→5'外切酶活性很低。相反,另外一些具有抗突变能力的T4突变株中的T4DNA聚合酶的3'→5'外切酶活性比野生型高得多,因此,其DNA复制真实性好,变异率低。可见,3'→5'外切酶活性对DNA复制真实性的维持是十分重要的。
[3]5'→3'外切酶活性──切除修复作用:5'→3'外切酶活性就是从5'→3'方向水解DNA生长链前方的DNA链,主要产生5'-脱氧核苷酸。这种酶活性只对DNA上配对部份(双链)磷酸二酯键有切割活力作用,方向是5'→3'。每次能切除10个核苷酸,而且DNA的聚合作用能刺激5'→3'外切酶活力达10倍以上。因此,这种酶活性在DNA损伤的修复中可能起着重要作用。对冈崎片段5'末端RNA引物的去除依赖此种外切酶活性。
[4]焦磷酸解作用:DNApolⅠ的这种活性可以催化3'末端焦磷酸解DNA分子。这种作用就是无机焦磷酸分解DNA生长链,可以认为是DNA聚合作用的逆反应,而且这种水解DNA链作用需要有模板DNA的存在。(dNMP)n+XPPi←(dNMP)n-x+X(dNPPP)→DNA
[5]焦磷酸交换作用:催化dNTP末端的PPi同无机焦磷酸的交换反应。反应式为32P32Pi+dNPPP←dNP32P32P+PPi→DNA
最后两种作用,都要求有较高浓度的PPi,因此,在体内由于没有足够高的PPi而无重要意义。
DNApolⅠ的DNA聚合酶活性和5'→3'外切酶活性协同作用,可以使DNA链上的切口向前推进,即没有新的DNA合成,只有核苷酸的交换。这种反应叫缺口平移(Nick translation)。当双链DNA上某个磷酸二酯键断裂产生切口时,DNApoIⅠ能从切口开始合成新的NDA链,同时切除原来的旧链。这样,从切口开始合成了一条与被取代的旧链完全相同的新链。如果新掺入的脱氧核苷酸三磷酸为α-32P-dNTP,则重新合成的新链即为带有同位素标记的DNA分子,可以用作探针进行分子杂交实验。
尽管DNApolⅠ是第一个被鉴定的DNA聚合酶,但它不是在肠杆菌中DNA复制的主要聚合酶。主要证据如下:[1]纯化的DNApolⅠ催化dNTP掺入的速率为667碱基/分,而体内DNA合成速率要比此高二倍数量级;[2]大肠杆菌的一个突变株中,此酶的活力正常,但染色体DNA复制不正常;[3]而在另一些突变株中,DNApolⅠ的活力中只是野生型的1%,但是DNA复制却正常,而且此突变株增加了对紫外线、烷化剂等突变因素的敏感性。这表明该酶与DNA复制关系不大,而在DNA修复中起着重要的作用。
大肠杆菌DNA聚合酶
1、大肠杆菌DNA聚合酶Ⅰ(DNApolymeraseⅠ,DNApolⅠ)这是1956年由ArthurKornberg首先发现的DNA聚合酶,又称Kornber酶。此酶研究得清楚而且代表了其他DNA聚合酶的基本特点。
(1)理化性质:纯化的DNApolⅠ由一条多肽链组成,约含1000个氨基酸残基,MW为109KD。分子含有一个二硫键和一个-SH基。通过二个酶分子上的-SH基与Hg2+结合产生二聚体,仍有活性。每个酶分子中含有一个Zn2+,在DNA聚合反应起着很重要的作用。每个大肠杆菌细胞中含有约400个分子,每个分子每分钟在37℃下能催化667个核苷酸掺入正在生长的DNA链。经过枯草杆菌蛋白酶处理后,酶分子分裂成两个片段,大片段分子量为76KD,通常称为Klenow片段,小片段的分子量为34KD。此酶的模板专一性和底物专一性均较差,它可以用人工合成的RNA作为模板,也可以用核苷酸为底物。在无模板和引物时还可以从头合成同聚物或异聚物。
DNAPolⅠ在空间结构上近似球体,直径约65A。在酶的纵轴上有一个约20A的深沟(cleft),带有正电荷,这是该酶的活性中心位置,在此位置上至少有6个结合位点:[1]模板DNA结合位点;[2]DNA生长链或引物结合位点;[3]引物末端结合位点,用以专一引物或DNA生长链的3'-OH;[4]脱氧核苷三磷酸结合位点;[5]5'→3'外切酶活性位点,用以结合生长链前方的5'-端脱氧核苷酸并切除之;[6]3'→5'外切酶活性位点,用以结合和切除生长链上未配对的3'-端核苷酸。
2、大肠杆菌DNA聚合酶Ⅱ(DNApolⅡ):DNA聚合酶Ⅰ缺陷的突变株仍能生存,这表明DNApolⅠ不是DNA复制的主要聚合酶。人们开始寻找另外的DNA聚合酶,并于1970年发现了DNApolⅡ。此酶MW为120KD,每个细胞内约有100个酶分子,但活性只有DNApolⅠ的5%。该酶的催化特性如下:
(1)聚合作用:该酶催化DNA的聚合,但是对模板有特殊的要求。该酶的最适模板是双链DNA而中间有空隙(gap)的单链DNA部份,而且该单链空隙部份不长于100个核苷酸。对于较长的单链DNA模板区该酶的聚合活性很低。但是用单链结合蛋白(SSBP)可以提高其聚合速率,可达原来的50-100倍。
(2)该酶也具有3'→5'外切酶活性,但无5'→3'外切酶活性。
(3)该酶对作用底物的选择性较强,一般只能将2-脱氧核苷酸掺入到DNA链中。
(4)该酶不是复制的主要聚合酶,因为此酶缺陷的大肠杆菌突变株的DNA复制都正常。可能在DNA的损伤修复中该酶起到一定的作用。
3、大肠杆菌DNA聚合酶Ⅲ(DNApolⅢ):DNA pol Ⅲ全酶由多种亚基组成(见下表),而且容易分解。大肠杆菌每个细胞中只有10-20个酶分子,因此不易获得纯品,为研究该酶的各种性质和功能带来了许多困难。直到不久前,才对其性质和功能有所了解,但每个亚基的具体作用扔不十分清楚。尽管该酶在细胞内存在的数量较少,但催化脱氧核苷酸掺入DNA链的速率分别是DNA聚合酶Ⅱ的15倍和30倍。该酶对模板的要求与DNA聚合酶Ⅱ相同,最适模板也是链DNA中间有空隙的单链DNA,单链结合蛋白可以提高该酶催化单链DNA模板的DNA聚合作用。DNApolⅢ也有3'→5'和5'→3'外切酶活性,但是3'→5'外切酶活性的最适底物是单链是DNA,只产生5'-单核苷酸,不会产生二核苷酸,即每次只能从3'端开始切除一个核苷酸。5'→3'外切酶活性也要求有单链DNA为起始作用底物,但一旦开始后,便可作用于双链区。DNA聚合酶Ⅲ是细胞内DNA复制所必需的酶,缺乏该酶的温度突变株在限制温度(non permissive temperature)内是不能生长的,此种突变株的裂解液也不能合DNA,但加入DNA聚合酶Ⅲ则可以恢复其合成DNA的能力。
DNA聚合酶Ⅲ的组成
亚基 分子量(×103) 其它名称
a 140 dnaE蛋白,polC蛋白
ε 25
θ 10
τ 83
γ 52 dnaZ蛋白
δ 32 dnaX蛋白
β 40 dnaN蛋白,copolⅢ
大肠杆菌三种DNA聚合酶的特性
功能 polⅠ polⅡ polⅢ
聚合作用5'→3' + +
外切酶活性3'→5' + + +
外切酶活性5'→3' + + +
焦磷酸解和焦磷酸交换作用 + - +
模板及引物的选择
完整的DNA双链 - - -
带引物的长单链DNA + - -
带缺口的双链DNA + - -
双链而有间障的DNA + + +
一般性质
分子量 1 09KD 120KD >140KD
每细胞中的分子量 400 17-100 10-20
结构基因 polA polB polC
T4-DNA聚合酶
T4-DNA聚合酶(T4-DNApol):近年来在枯草杆菌,鼠伤寒沙门氏杆菌等细胞及噬菌体T4、T5、T7中分离到NDA聚合酶,这里只介绍T4DNA聚合酶。T4DNA聚合酶与大肠杆菌DNA聚合酶Ⅰ相似,也是一条多肽链,分子量亦相近,但氨基酸组成不同,它至少含有15个半胱氨酸残基。但是,它在作用上与大肠杆菌DNApol不同;[1]它无5'→3'外切酶活性;[2]它需要一条有引物的单链DNA作模板。在有缺口的双链DNA作模板时,需要有基因32蛋白的辅助(基因32蛋白在T4DNA复制中的作用和大肠杆菌单链结合蛋白的作用相似,基因32蛋白在复制叉处和单链DNA结合后可以促进双链的进一步打开,并保持其单链状态有利于新生链的合成);它利用单链DNA为模板进,可同时利用它作为引物,即此单链DNA的3'端能环绕其本身的某一顺序形成氢键配对,3'端的未杂交部分即被T4DNA聚合酶的3'→5'外切酶活性切去,然后在其作用下从3'-OH端开始聚合,合成该模板DNA的互补链,再以互补链为模板合成原来的单链DNA。
真核生物的DNA聚合酶
真核生物的DNA聚合酶:真核生物中也具有几种DNA聚合酶,但这些聚合酶都没有3'→5'或5'→3'外切酶活性。其聚合反应机制与原核生物的聚合一样。主要的酶(占总量的80-90%)是DNA聚合酶α,分子量为300KD,含有4个或5个亚基,主要负责染色体DNA的复制。DNA聚合酶β,分子量为45KD,仅含有一条链,主要作用是修复核内DNA。第三种聚合酶γ,分子量为140KD,存在于线粒体内,负责催化线粒体DNA的复制。最近从兔的骨髓细胞质中分离到一种新的DNA聚合酶,这种酶与上述真核生物的DNA聚合酶不同,而与原核生物的聚合酶类似,具有3'→5'核酸外切酶活性,称为DNA聚合酶δ。另外,从酵母、原虫等低真核生物中分离到一些DNA聚合酶。一般来说,这些低等生物都没有低分子量的DNA聚合酶β,而且与原核生物的DNA聚合酶相似,有3'→5'外切酶活性。
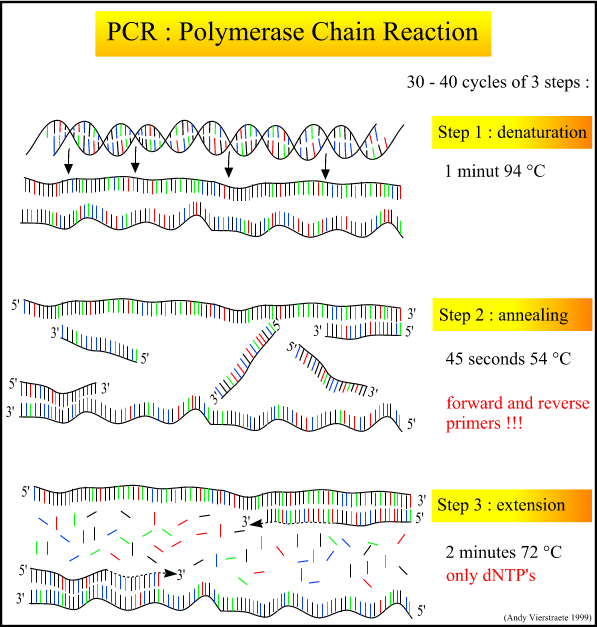
实验室常用的耐热DNA聚合酶
耐热DNA聚合酶多应用在PCR技术中。各种耐热DNA聚合酶均具有5'-3'聚合酶活性,但不一定具有3'-5'和5'-3'的外切酶活性。3'-5'外切酶活性可以消除错配,切平末端;5'-3'外切酶活性可以消除合成障碍。由于上述这些不同,可以将耐热DNA聚合酶分为三类:
一、普通耐热DNA聚合酶
Taq DNA 聚合酶
由一种水生栖热菌yT1株分离提取出,是发现的耐热DNA聚合酶中活性最高的一种,达200,000单位/mg。具有5'-3'外切酶活性,但不具有3'-5'外切酶活性,因而在合成中对某些单核苷酸错配没有校正功能。
TaqDNA聚合酶还要具有非模板依赖性活性,可将PCR双链产物的每一条链3'加入单核苷酸尾,故可使PCR产物具有3'突出的单A核苷酸尾;另一方面,在仅有dTTP存在时,它可将平端的质粒的3'端加入单T核苷酸尾,产生3'端突出的单T核苷酸尾。应用这一特性,可实现PCR产物的T-A克隆法。
Tth DNA 聚合酶
从Thermus thermophilus HB8中提取而得,改酶在高温和MnCl2条件下,能有效地逆转录RNA;当加入Mg2+后,该酶的聚合活性大大增加,从而使cDNA合成与扩增可用一种酶催化。
二、高保真DNA聚合酶
pfu DNA 聚合酶
是从Pyrococcus furiosis中精制而成的高保真耐高温DNA聚合酶,它不具有5'-3'外切酶活性,但具有3'-5'外切酶活性,可校正PCR扩增过程中产生的错误,使产物的碱基错配率极低。PCR产物为平端,无3'端突出的单A核苷酸。
Vent DNA 聚合酶
改酶是从Litoralis栖热球菌中分离出的,不具有5'-3'外切酶活性,但具有3'-5'外切酶活性,可以去除错配的碱基,具有校对功能。
三、DNA序列测定中应用的耐热DNA聚合酶
Promega公司测序级TaqDNA聚合酶
是在TaqDNA聚合酶的基础上对它进行修饰,去除5'-3'外切酶活性,保证测序记过的高度准确性,可产生强度均一的测序条带,背景清晰。
Bca Best DNA 聚合酶
从Bacillus Caldotenax YT-G菌株中提纯,并使其5'-3'外切酶活性缺失的DNA聚合酶。它的伸长性能优越;可抑制DNA二级结构形成,可以得到均一的DNA测序带。
Sac DNA 聚合酶
从酸热浴流化裂片菌中分离,无3'-5'外切酶活性,可用于DNA测序,但测序反应中ddNTP/dNTP的比率要高。