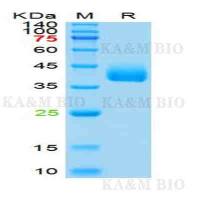
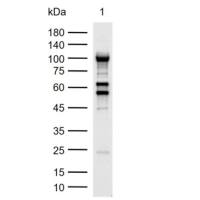
PCR技术:个体配子DNA序列的PCR分析
互联网
最近发展了一种不依赖遗传交叉或家系分析的方法检测遗传标记间的重组频率。 具体方法是直接决定配子(精子)的基因型是亲本型还是重组型。这通过确定两个等位 基因中哪一个存在于双杂合男性精子上的两个基因座点其中之一来完成。根据上述方 法确定的重组精子的数量被总检测精子数除就可估计出重组频率。已知聚合酶反应在 体外扩增基因能够使DNA扩增10 9 倍,因此该技术有可能分析精子中单分子靶DNA。以 这种方式研究数千个人精子就有可能构建比家系分析要精确得多的遗传图谱。该方法 提供了一个独物的手段来研究许多现在被认为是不能解决的人类遗传学领域的难题。
用PCR确定精子类型的原理
从人类家系分析的观点来看,每个个体都是亲本减数分裂产生的单个产物的融合 体。双亲以及后代体细胞DNA的研究加上适当的统计学方法可推知形成合子的减数分 裂产物的重组或非重组的性质,同时可以估计重组在减数分裂中所占的比例。我们研 究遗传重组的方法与上述的不同,它通过分析减数分裂产物本身的DNA来决定减数分 裂发生重组的比率。每个男性一生中可产生大量的减数分裂产物精子,从而提供了进 行遗传分析取之不尽的实验材料。
假设一个男性杂合子有两个连锁基因座。每一个基因座的多态性差异在于AT-AT 或GC-GC)要么是重组型(AT-GC或GC-AT)。重组类型出现的频率将取决于两个基因座彼 此间的遗传间距。通过检测某一个体的大量精子,可精确地计算出四种精子类型出现 的频率。该技术存在的困难在于确定减数分裂前细胞中的每个基因座上存在的两种等 位基因中的哪一个进入单个精子。常规的分子生物学技术没有足够高的灵敏度分析精 子染色体DNA的单个分子。
PCR扩增每个基因座的多态性区域是分析精子的第一步。两个基因座用两组引 物,同时引物必须位于多态性区域的边侧。当精子两个靶序列经PCR扩增,就确定了 每个基因座的等位基因成份。人工合成的寡核苷酸探针可识别等位基因小到仅发生单 个碱基置换的实验方法已得到改进,这些等位基因牧场划的寡聚物(ASO)探针第一 次应用是将人正常β珠蛋白基因中的正常βA等位基因与镰刀形红细胞(βA)的突变 区分开来。本方法检测等位基因需要人工合成的小片段寡核苷酸(典型的长19bp)。 每个寡核苷酸与一个等位基因完全互补而与另一个等位基因通常只有一个碱基的差 别。同位素标记的寡核苷酸分别与待检测扩增的DNA样品杂交。在适当的杂交条件 下,只有样品中的序列完全互补时,寡核苷酸可形成稳定的双螺旋。例如,精子的 PCR产物用四种ASO探针分析。有一对ASO分别将基因座1的两个等位基因区别开,另一 对将基因座2的两个等位基因区别开。实际上,预期两个ASO中只有一个与某一个减数 分裂产物杂交,可以确定每个检测的精子对于所研究的染色体而言实际上是单倍体。
人单倍体细胞有1.5-5X10 -24 摩尔单一DNA序列。根据以前的实验结果,我们知道 活性达5μCI/微微摩尔的同位素标记就能够在不到一天的时间内检测出1毫微微摩尔 (10 -15 /10 -24 )。即使以较低的平均扩增效率,大约经50个扩增循环便可完成。