细胞自噬系统介绍
互联网
15012
自噬是近年来很热门的领域,做一下系统的介绍或讨论,内容:
1) 什么是自噬?
包括自噬的定义、形态学特征、分子基础、调控等
2)自噬的意义
自噬与细胞存活、细胞死亡、疾病、衰老等的关系
3)怎么研究自噬?
主要谈谈怎么证明细胞发生了自噬,即自噬的判断标准和各种研究自噬的方法及局限性。
一、自噬的过程——从一张图片开始
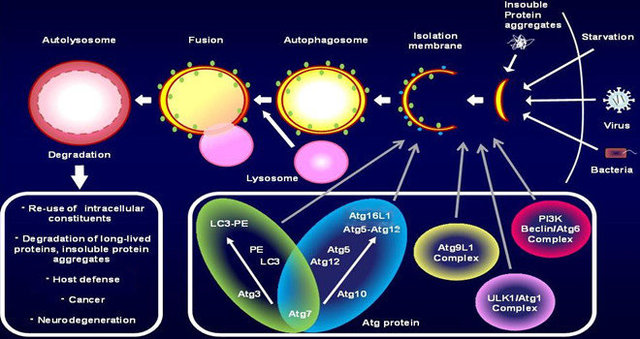
步骤1:细胞接受自噬诱导信号后,在胞浆的某处形成一个小的类似“脂质体”样的膜结构,然后不断扩张,但它并不呈球形,而是扁平的,就像一个由2层脂双层组成的碗,可在电镜下观察到,被称为Phagophore,是自噬发生的铁证之一。
步骤2:Phagophore不断延伸,将胞浆中的任何成分,包括细胞器,全部揽入“碗”中,然后“收口”,成为密闭的球状的autophagosome,我把它翻译为“自噬体”。电镜下观察到自噬体是自噬发生的铁证之二。有2个特征:一是双层膜,二是内含胞浆成分,如线粒体、内质网碎片等。
步骤3:自噬体形成后,可与细胞内吞的吞噬泡、吞饮泡和内体融合(加了个“可”字,意思是这种情况不是必然要发生的)。
步骤4:自噬体与溶酶体融合形成autolysosome,期间自噬体的内膜被溶酶体酶降解,2者的内容物合为一体,自噬体中的“货物”也被降解,产物(氨基酸、脂肪酸等)被输送到胞浆中,供细胞重新利用,而残渣或被排出细胞外或滞留在胞浆中。

最详细的介绍:Madame Curie Bioscience Database
http://www.ncbi.nlm.nih.gov/books/bv.fcgi?rid=eurekah.chapter.24952
透射电镜下自噬的照片:

上图的英文说明:

二、自噬的特性
1)自噬是细胞消化掉自身的一部分,即self-eating,初一看似乎对细胞不利。事实上,细胞正常情况下很少发生自噬,除非有诱发因素的存在。这些诱发因素很多,也是研究的热门。既有来自于细胞外的(如外界中的营养成分、缺血缺氧、生长因子的浓度等),也有细胞内的(代谢压力、衰老或破损的细胞器、折叠错误或聚集的蛋白质等)。由于这些因素的经常性存在,因此,细胞保持了一种很低的、基础的自噬活性以维持自稳。
2)自噬过程很快,被诱导后8min即可观察到自噬体(autophagosome)形成,2h后自噬溶酶体(autolysosome)基本降解消失。这有利于细胞快速适应恶劣环境。
3)自噬的可诱导特性:表现在2个方面,第一是自噬相关蛋白的快速合成,这是准备阶段。第二是自噬体的快速大量形成,这是执行阶段。
4)批量降解:这是与蛋白酶体降解途径的显著区别。
5)“捕获”胞浆成分的非特异性:由于自噬的速度要快、量要大,因此特异性不是首先考虑的,这与自噬的应急特性是相适应的。
6)自噬的保守性:由于自噬有利于细胞的存活,因此无论是物种间、还是各细胞类型之间(包括肿瘤细胞),自噬都普遍被保留下来(谁不喜欢留一手呢?)。
参考综述:
Autophagy fights disease through cellular self-digestion.pdf
http://www.namipan.com/d/4c78fe31cb24080cd7630639c121cd71b24c9b56fbdf6300
自噬相关基因(autophagy associated gene, ATG):在自噬过程中到底有哪些蛋白的参与,即自噬相关蛋白的鉴定是目前自噬研究主要的任务。由于自噬研究的历史关系,很多基因在酵母和哺乳动物中有不同的命名。
在http://www.ncbi.nlm.nih.gov/books/bv.fcgi?rid=eurekah.table.24964中介绍了一些较早发现的自噬相关基因,下面列出几个新近发现的:
1)bif-1(又叫Endophilin B1) 和 UVRAG(ultraviolet irradiation resistance-associated gene)
相关文章:Bif-1 interacts with Beclin 1 through UVRAG and regulates autophagy and tumorigenesis.pdf
http://www.namipan.com/d/75c245aa6c8cf8644c33bcf86e8c6ef8add3da69469e0e00
2)VMP1 (Vacuole membrane protein 1)
相关文章:The Pancreatitis-induced Vacuole Membrane Protein 1 Triggers Autophagy in Mammalian Cells.pdf
http://www.namipan.com/d/a4a74e47a1d515089558fe39db2177ff931ee8114a611e00
3)DRAM (damage-regulated autophagy modulator)
相关文章:DRAM, a p53-Induced Modulator of Autophagy, Is Critical for Apoptosis.pdf
http://www.namipan.com/d/69fee934467a3fc4b9a70b88c9292160a631c78f42111300
4)TP53INP2 (Tumour Protein 53 Induced Nuclear Protein 2)
相关文章:The TP53INP2 Protein Is Required for Autophagy in Mammalian Cells.pdf
http://www.namipan.com/d/40e5d3a2f6a0ec5fce4dc91c32880ae293201a582cf33500
(由于不断有新的ATG蛋白被鉴定出来,本帖将不定期更新)
三、自噬过程的调控
从上面总结的自噬特点中可以看出,自噬这一过程一旦启动,必须在度过危机后适时停止,否则,其非特异性捕获胞浆成分的特性将导致细胞发生不可逆的损伤。这也提醒我们在研究自噬时一定要动态观察,任何横断面的研究结果都不足以评价自噬的活性。
目前,已经报告了很多因素能诱导细胞发生自噬,如饥饿、生长因子缺乏、微生物感染、细胞器损伤、蛋白质折叠错误或聚集、DNA损伤、放疗、化疗等等,这么多刺激信号如何传递的、哪些自噬蛋白接受信号、又有哪些自噬蛋白去执行等很多问题都还在等待进一步解答中。
关于传递自噬信号的通路目前比较肯定的有:
抑制类
1)Class I PI3K pathway (PI——phosphatidylinositol,磷脂酰肌醇)
与IRS (Insulin receptor substrate) 结合,接受胰岛素受体传来的信号(血糖水平高抑制自噬)
详细介绍参见:
维基百科 http://en.wikipedia.org/wiki/Phosphoinositide_3-kinase
http://www.sigmaaldrich.com/life-science/cell-biology/learning-center/pathfinder/pathway-maps/pi3k-signaling.html
2)mTOR pathway(mammalian target of rapamycin)
mTOR在人类中的同源基因是FRAP1(FK506 binding protein 12-rapamycin associated protein 1),是一个丝/苏氨酸蛋白激酶。能接受多种上游信号,如Class I PI3K、IGF-1/2、MAPK,能感受营养和能量的变化。
详细介绍见:
http://en.wikipedia.org/wiki/MTOR
激活类
1)Class III PI3K
结构上类似于Class I PI3K,但作用相反。
接受上述信号的自噬蛋白:
目前都把焦点集中在beclin 1(酵母同源物为atg6),能与多种蛋白结合,如Vps34(Class III PI3K的催化亚单位),mTOR,BCL-2和BCLXL蛋白等,更多介绍请参阅OMIM:http://www.ncbi.nlm.nih.gov/entrez/dispomim.cgi?id=604378。
在【动态版】http://www.dxy.cn/bbs/post/view?bid=154&id=14115812&sty=1中介绍了日本科学家最新的发现:beclin-1有可能成为人工干预自噬活性的靶点。
需注意的是,beclin-1是一个多功能蛋白,除了接受自噬信号,它还可以接受很多其它的信号对自噬进行调节,越来越多的证据表明,beclin-1可能是自噬的“守门人”。
四、自噬体的发生
目前认为,自噬体的膜不是直接来源于高尔基体或内质网,而是在胞浆中重新生成的,但具体的机制尚不清楚。
当beclin-1被活化后,胞浆中先形成很多个membrane source(自噬体膜发生中心),在它们不断扩展的过程中(phagophore到autolysosome),VMP1蛋白由内质网和高尔基体转位到自噬体膜上(VMP1又叫TMEM49,已知唯一与自噬有关的跨膜蛋白),同时,MAP1-LC3由胞浆型(即LC3-I)转位到自噬体膜(即LC3-II),LC3这一转变过程可被Western Blot和荧光显微镜检测到,现已成为监测自噬体形成的推荐方法。
五、自噬与细胞死亡的关系
有必要说明一下的是,细胞死亡是一个非常复杂的过程,为了研究方便,需进行分类,但我们思考时不要局限于这些人为的分类,而应注重于现象本身来研究其背后的机制。
一直以来人们从不同角度、用不同方法来观察细胞的死亡,并把细胞的死亡方式分为2类:坏死和凋亡,因为两者有着明显的区别,其中最主要的区别之一就是细胞膜的通透性——坏死细胞的细胞膜丧失了完整性,内容物被释放出来,染料可自由进入细胞,而凋亡细胞保持完整,无内容物释放,染料也被排斥。很多实验亦根据这一原理来设计以区分坏死和凋亡,这将在后面一一介绍,如同刚刚说明的那样,这些实验只能说明细胞膜的通透性(必要条件,不是充分必要条件),而不能用来证实坏死细胞或凋亡细胞。
一般认为坏死是被动的,不可控的,而凋亡是主动的,可控的。为了强调这一点,凋亡被定义为程序性细胞死亡(program cell death,PCD)。但无论是坏死还是凋亡,都是一个过程,是需要时间的(尤其是凋亡,从启动到完成,细胞要执行很多反应),而且细胞死亡后都有“尸体”。
在研究自噬与凋亡的关系时,人们发现细胞死亡前胞浆中存在大量的自噬体或自噬溶酶体,但这样的细胞缺乏凋亡的典型特点,如核固缩(pyknosis), 核破裂(karyorhexis)、细胞皱缩(shrinkage)、没有凋亡小体的形成等,被称为自噬样细胞死亡(autophagic cell death,ACD),它是一种新的细胞程序性死亡,为了与凋亡区别,被命名为Type II cell death,相应的,凋亡为Type I cell death,坏死为Type III cell death。
尽管这样,但对于自噬是否是细胞死亡的直接原因目前还存在很大的争议。到底是Cell death by autophagy(自噬引起死亡)还是Cell death with autophagy(死亡时有自噬发生,但不是直接原因)?
对此,自噬研究领域“大牛”级专家Levine Beth在一篇nature的Review中表达了自己的观点。
由于在形态学上2者无明显区别,但通过阻断自噬,观察细胞的结局可区分开来:Cell death by autophagy细胞存活,而Cell death with autophagy细胞死亡。
Autophagic cell death: the story of a misnomer.pdf
六、自噬与肿瘤的关系
与凋亡(在肿瘤细胞中一般都存在缺限)不同,自噬是被优先保留的。无论是肿瘤细胞还是正常细胞,保持一种基础、低水平的自噬活性是至关重要的。因为细胞中随时产生的“垃圾”(破损或衰老的细胞器、长寿命蛋白质、错误合成或折叠错误的蛋白质等等)都需要及时清除,而这主要靠自噬来完成,因此,自噬具有维持细胞自稳的功能;如果将自噬相关基因突变失活,如神经元会发生大量聚集蛋白,并出现神经元退化。同时,自噬的产物,如氨基酸、脂肪酸等小分子物质又可为细胞提供一定的能量和合成底物,可以说,自噬就是一个“备用仓库”。如Atg-5缺陷的小鼠在出生后喝上第一口奶之前就会饿死。更重要的是,自噬活性可在代谢应激(饥饿、生长因子缺乏、射线、化疗等)时大大增强,表现为胞浆中迅速涌现大量自噬体,这一现象被称为“自噬潮”(autophagic flux),广泛用于自噬形成的监测。自噬潮为细胞度过危机提供了紧急的营养和能量支持,有利于细胞的存活。
鉴于自噬的上述作用,自噬可为肿瘤细胞带来几大好处:
1)肿瘤细胞本身就具有高代谢的特点,对营养和能量的需求比正常细胞更高,但肿瘤微环境往往不能如意,如肿瘤发生初始期到血管发生之前、肿瘤长大发生血管崩塌时、肿瘤细胞脱离原发灶游走时等都会出现营养不足或供应中断,而此时提高自噬活性可以有助于度过这一危机。
2)当化疗、放疗后,肿瘤细胞会产生大量的破损细胞器、损坏的蛋白质等有害成分,而此时提高自噬活性可及时清除这些有害物质,并提供应急的底物和能量为修复受损DNA赢得时间和条件。
由于自噬减少了肿瘤细胞在代谢应激时发生坏死的机会,而对于肿瘤细胞群体而言,需要一部分细胞发生坏死,以引发适度的炎症(有利于血管的长入、吸引免疫细胞分泌生长因子等)。研究发现,很多类型的肿瘤在代谢应激时会“组成性”活化PI3K信号以抑制自噬(由于凋亡通路已受阻,抑制自噬会促进坏死),但具体机制尚不清楚。
参考文献:
Autophagy and toll-like receptors:A new link in cancer cells.pdf
Autophagy fights disease through cellular self-digestion.pdf
Role and regulation of autophagy in cancer.pdf
Role of autophagy in cancer.pdf
Autophagy and Cell Death: No Longer at Odds.pdf
Autophagy and cancer: Dynamism of the metabolism of tumor cells and tissues.pdf
七、自噬的研究方法
正常培养的细胞自噬活性很低,不适于观察,因此,必须对自噬进行人工干预和调节,经报道的工具药有:
(一)自噬诱导剂
1)Bredeldin A / Thapsigargin / Tunicamycin :模拟内质网应激
2)Carbamazepine/ L-690,330/ Lithium Chloride(氯化锂):IMPase 抑制剂(即Inositol monophosphatase,肌醇单磷酸酶)
3)Earle's平衡盐溶液:制造饥饿
4)N-Acetyl-D-sphingosine(C2-ceramide):Class I PI3K Pathway抑制剂
5)Rapamycin:mTOR抑制剂
6)Xestospongin B/C:IP3R阻滞剂
(二)自噬抑制剂
1)3-Methyladenine(3-MA):(Class III PI3K) hVps34 抑制剂
2)Bafilomycin A1:质子泵抑制剂
3)Hydroxychloroquine(羟氯喹):Lysosomal lumen alkalizer(溶酶体腔碱化剂)
除了选用上述工具药外,一般还需结合遗传学技术对自噬相关基因进行干预:包括反义RNA干扰技术(Knockdown)、突变株筛选、外源基因导入等。
细胞经诱导或抑制后,需对自噬过程进行观察和检测,常用的策略和技术有:
1)观察自噬体的形成
由于自噬体属于亚细胞结构,普通光镜下看不到,因此,直接观察自噬体需在透射电镜下。Phagophore的特征为:新月状或杯状,双层或多层膜,有包绕胞浆成分的趋势。自噬体(AV1)的特征为:双层或多层膜的液泡状结构,内含胞浆成分,如线粒体、内质网、核糖体等。自噬溶酶体(AV2)的特征为:单层膜,胞浆成分已降解。(autophagic vacuole,AV)
2)在荧光显微镜下采用GFP-LC3融合蛋白来示踪自噬形成:
由于电镜耗时长,不利于监测(Monitoring)自噬形成,人们利用LC3在自噬形成过程中发生聚集的现象开发出了此技术。无自噬时,GFP-LC3融合蛋白弥散在胞浆中;自噬形成时,GFP-LC3融合蛋白转位至自噬体膜,在荧光显微镜下形成多个明亮的绿色荧光斑点,一个斑点相当于一个自噬体,可以通过计数来评价自噬活性的高低。
3)利用Western Blot检测LC3-II/I比值的变化以评价自噬形成:
自噬形成时,胞浆型LC3(即LC3-I)会酶解掉一小段多肽,转变为(自噬体)膜型(即LC3-II),因此,LC3-II/I比值的大小可估计自噬水平的高低(Note:LC3抗体对LC3-II有更高的亲和力,会造成假阳性。方法2和3需结合使用,同时需考虑溶酶体活性的影响。)
4)检测长寿蛋白的批量降解:非特异
5)MDC(Monodansylcadaverine,单丹磺酰尸胺)染色:包括自噬体,所有酸性液泡都被染色,故属于非特异性的。
6)CellTrackerTM Green染色:主要用于双染色,但其能染所有的液泡,故也属于非特异性的。
在研究自噬相关蛋白时,需对其进行定位。由于自噬体与溶酶体、线粒体、内质网、高尔基体关系密切,为了区别,常用到一些示踪蛋白在荧光显微镜下来共定位:
Lamp-2:溶酶体膜蛋白,可用于监测自噬体与溶酶体融合。
LysoTrackerTM 探针:有红或蓝色可选,显示所有酸性液泡。
pDsRed2-mito:载体,转染后表达一个融合蛋白(红色荧光蛋白+线粒体基质定位信号),可用来检测线粒体被自噬掉的程度(Mitophagy)。
MitoTraker探针:特异性显示活的线粒体,荧光在经过固定后还能保留。
Hsp60:定位与线粒体基质,细胞死亡时不会被释放。
Calreticulin(钙网织蛋白):内质网腔
【Note:这些蛋白均为胞浆蛋白,爬片或胰酶消化的细胞在做免疫荧光前需先透膜(permeablize),可采用0.1%SDS处理】
自噬与细胞死亡经常需一起考虑,下面介绍一些检测细胞死亡的方法:
(1)△ψm dissipation(线粒体跨膜电位的消失):TMRM发红色荧光,DiOC6(3)发绿色荧光。
(2)Phosphatidylserine Externalization(磷脂酰丝氨酸外翻):Annexin V-FITC(绿色)染细胞膜,(3)检测线粒体产生的ROS:无荧光的HE(hydroethidine,氢化乙啶)可被ROS氧化为EthBr(ethidium bromide,溴乙啡啶),发红色荧光。NAO(nonyl acridine orange,烷化吖啶橙,可发荧光)能与非氧化的cardiolipin(心磷脂,可被ROS氧化)反应而失去荧光。
(4)线粒体IMS蛋白的释放:AIF,细胞色素c,分别用荧光二抗染色。
(5)Capase 3a 染色:用荧光二抗染色,胞浆弥散分布。
(6)细胞膜完整性检测:DAPI(蓝色)、Hoechst 33342或PI(红色)染核。胞膜完整的细胞(活细胞和早中期凋亡细胞)排斥,可联用annexin V。
【如何用实验区分Cell death by autophagy和Cell death with autophagy】
第一步:利用上述方法证实细胞死亡
第二步:证实细胞死亡前发生了自噬
第三步:在形态学上区别开“自噬样死亡”与凋亡
第四步:利用遗传学手段(反义RNA干扰Knockdown掉Atg基因或hVps34)或工具药抑制自噬
第五步:细胞存活则为Cell death by autophagy,反之,细胞死亡则为Cell death with autophagy。
参考文献:
Methods for Assessing Autophagy and Autophagic Cell Death.pdf
Cytosolic LC3 Ratio as a Sensitive Index of Macroautophagy in Isolated Rat Hepatocytes and H4-II-E Cells.pdf
讨论贴:http://www.dxy.cn/bbs/topic/14167726
讨论贴:http://www.dxy.cn/bbs/topic/14167726