噬菌体展示技术
互联网
噬菌体展示技术是将外源蛋白或多肽的DNA序列插入到噬菌体外壳蛋白结构基因的适当位置,使外源基因随外壳蛋白的表达而表达,同时,外源蛋白随噬菌体的重新组装而展示到噬菌体表面的生物技术。到目前为止,人们已开发出了单链丝状噬菌体展示系统、λ噬菌体展示系统、T4噬菌体展示系统等数种噬菌体展示系统。本文主要概述了噬菌体展示技术的基本原理、噬菌体展示系统研究以及技术特点等,并跟踪了目前该领域的最新研究进展和发展前景。
关键词:噬菌体展示;组装;融和蛋白
1985年,Smith G P[1]第一次将外源基因插入丝状噬菌体f1的基因Ⅲ,使目的基因编码的多肽以融合蛋白的形式展示在噬菌体表面,从而创建了噬菌体展示技术。该技术的主要特点是将特定分子的基因型和表型统一在同一病毒颗粒内,即在噬菌体表面展示特定蛋白质,而在噬菌体核心DNA中则含有该蛋白的结构基因。
另外,这项技术把基因表达产物与亲和筛选结合起来,可以利用适当的靶蛋白将目的蛋白或多肽挑选出来。近年来,随着噬菌体展示技术的日益完善,该技术在众多基础和应用研究领域产生的影响已日渐明显。
一、 噬菌体展示技术的原理
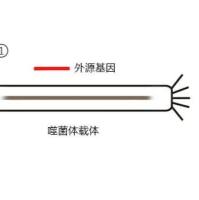
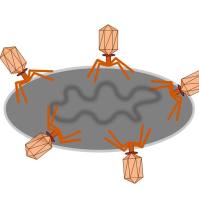
噬菌体展示技术是将多肽或蛋白质的编码基因或目的基因片段克隆入噬菌体外壳蛋白结构基因的适当位置,在阅读框正确且不影响其他外壳蛋白正常功能的情况下,使外源多肽或蛋白与外壳蛋白融合表达,融合蛋白随子代噬菌体的重新组装而展示在噬菌体表面。被展示的多肽或蛋白可以保持相对独立的空间结构和生物活性,以利于靶分子的识别和结合。肽库与固相上的靶蛋白分子经过一定时间孵育后,洗去未结合的游离噬菌体,然后以竞争受体或酸洗脱下与靶分子结合吸附的噬菌体,洗脱的噬菌体感染宿主细胞后经繁殖扩增,进行下一轮洗脱,经过3轮~5轮的“吸附-洗脱-扩增”后,与靶分子特异结合的噬菌体得到高度富集[2]。所得的噬菌体制剂可用来做进一步富集有期望结合特性的目标噬菌体。
二、 噬菌体展示系统
2.1 单链丝状噬菌体展示系统
(1)PⅢ展示系统。
丝状噬菌体是单链DNA病毒,PⅢ是病毒的次要外壳蛋白,位于病毒颗粒的尾端,是噬菌体感染大肠埃希菌所必须的。每个病毒颗粒都有3个~5个拷贝PⅢ蛋白[3],其在结构上可分为N1、N2和CT 3个功能区域[4-5],这3个功能区域由两段富含甘氨酸的连接肽G1和G2连接。其中,N1和N2与噬菌体吸附大肠埃希菌菌毛及穿透细胞膜有关[6],而CT构成噬菌体外壳蛋白结构的一部分,并将整个PⅢ蛋白的C端结构域锚定于噬菌体的一端[7]。PⅢ有2个位点可供外源序列插入,当外源的多肽或蛋白质融合于PⅢ蛋白的信号肽(SgⅢ)和N1之间时,该系统保留了完整的PⅢ蛋白,噬菌体仍有感染性;但若外源多肽或蛋白直接与PⅢ蛋白的CT结构域相连,则噬菌体丧失感染性,这时重组噬菌体的感染性由辅助噬菌体表达的完整PⅢ蛋白来提供。PⅢ蛋白很容易被蛋白水解酶水解,所以有辅助噬菌体超感染时,可以使每个噬菌体平均展示不到一个融合蛋白,即所谓“单价”噬菌体。
(2)PⅧ及其他展示系统。
PⅧ是丝状噬菌体的主要外壳蛋白,位于噬菌体外侧,C端与DNA结合,N端伸出噬菌体外,每个病毒颗粒有2 700个左右PⅧ拷贝[8-9]。PⅧ的N端附近可融合五肽,但不能融合更长的肽链,因为较大的多肽或蛋白会造成空间障碍,影响噬菌体装配[2,10-11],使其失去感染力。但有辅助噬菌体参与时,可提供野生型PⅧ蛋白,降低价数,此时可融合多肽甚至抗体片段。 此外,尚有丝状噬菌体PⅥ展示系统的研究报道[12]。PⅥ蛋白的C端暴露于噬菌体表面,可以作为外源蛋白的融合位点,可以用于研究外源蛋白C端结构区域功能。从所掌握的文献来看,该系统主要用于cDNA表面展示文库的构建,并取得了不错的筛选效果。
2.2 λ噬菌体展示系统
(1)PV展示系统。
λ噬菌体的PV蛋白构成了它的尾部管状部分,该管状结构由32个盘状结构组成,每个盘又由6个PV亚基组成。PV有两个折叠区域,C端的折叠结构域(非功能区)可供外源序列插入或替换。目前,用PV系统已成功展示了有活性的大分子蛋白β-半乳糖苷酶(465 ku)和植物外源凝血素BPA(120 ku)等[13]。λ噬菌体的装配在细胞内进行,故可以展示难以分泌的肽或蛋白质。该系统展示的外源蛋白质的拷贝数为平均1个分子/噬菌体,这表明外源蛋白质或多肽可能干扰了λ噬菌体的尾部装配。
(2)D蛋白展示系统。
D蛋白的分子质量为11 ku,参与野生型λ噬菌体头部的装配。低温电镜分析表明,D蛋白以三聚体的形式突出在壳粒表面[13]。当突变型噬菌体基因组小于野生型基因组的82%时,可以在缺少D蛋白的情况下完成组装,故D蛋白可作为外源序列融合的载体,而且展示的外源多肽在空间上是可以接近的。病毒颗粒的组装可以在体内也可以在体外,体外组装即是将D融合蛋白结合到λD-噬菌体表面,而体内组装是将含D融合基因的质粒转化入λD-溶源的大肠埃希菌菌种中,从而补偿溶源菌所缺的D蛋白,通过热诱导而组装。该系统有一个很好的特点,噬菌体上融合蛋白和D蛋白的比例可以由宿主的抑制tRNA活性加以控制,这对于展示那些可以对噬菌体装配造成损害的蛋白质时特别有用。
2.3 T4噬菌体展示系统
T4噬菌体展示系统是20世纪90年代中期建立起来的一种新的展示系统。它的显著特点是能够将两种性质完全不同的外源多肽或蛋白质,分别与T4衣壳表面上的外壳蛋白SOC(9 ku)和HOC(40 ku)融合而直接展示于T4噬菌体的表面,因此它表达的蛋白不需要复杂的蛋白纯化,避免了因纯化而引起的蛋白质变性和丢失。T4噬菌体是在宿主细胞内装配,不需通过分泌途径,因而可展示各种大小的多肽或蛋白质,很少受到限制。吴健敏等成功地将大小约215 aa SOC/m E2融合蛋白展示于T4噬菌体衣壳表面[14]。令人值得关注的是,SOC与HOC蛋白的存在与否,并不影响T4的生存和繁殖。SOC和HOC在噬菌体组装时可优于DNA的包装而装配于衣壳的表面,事实上,在DNA包装被抑制时,T4是双股DNA噬菌体中唯一能够在体内产生空衣壳的噬菌体(SOC和HOC也同时组装)。因此,在用重组T4做疫苗时,它能在空衣壳表面展示目的抗原,这种缺乏DNA的空衣壳苗,在生物安全性方面具有十分光明的前景[14]。
三、 噬菌体展示技术的局限性
(1)在噬菌体展示过程中必须经过细菌转化、噬菌体包装,有的展示系统还要经过跨膜分泌过程,这就大大限制了所建库的容量和分子多样性。目前,常用的噬菌体展示文库中含有不同序列分子的数量一般限制在109。
(2)不是所有的序列都能在噬菌体中获得很好的表达,因为有些蛋白质功能的实现需要折叠、转运、膜插入和络合,导致在体内筛选时需外加选择压力。例如,在噬菌体展示文库试验中,由于部分未折叠的蛋白在细菌中很容易被降解,因此,必须小心控制条件,以保证在噬菌体表面展示的文库没有降解。另外,鼠源抗体在噬菌体中表达差,也是体内选择压力的一个例子。真核细胞蛋白在细菌中表达差是因为它们的蛋白质合成与折叠机制不同的缘故[15]。
(3)噬菌体展示文库一旦建成,很难再进行有效的体外突变和重组,进而限制了文库中分子遗传的多样性。
(4)由于噬菌体展示系统依赖于细胞内基因的表达,所以,一些对细胞有毒性的分子如生物毒素分子,很难得到有效表达和展示。
四、结语
噬菌体展示技术经过近20年的发展和完善,已成为生命科学领域的一项重要技术,广泛应用于抗原抗体库的建立、药物设计、疫苗研究、病原检测、基因治疗、抗原表位研究及细胞信号转导研究等[16]。噬菌体展示系统模拟了自然免疫系统,使我们有可能模拟体内抗体生成过程,构建高亲和力抗体库。由于噬菌体展示技术实现了基因型和表型的有效转换,使研究者在基因分子克隆基础上实现了蛋白质构象体外控制,从而为获取具有良好生物学活性的表达产物提供了强有力手段。另外,噬菌体展示技术已成为不经过免疫获取特异性人源抗体的新途径,为获取对人类和动物疾病有诊断和治疗价值的单克隆抗体提供了重要手段。