用RNaseIII制备siRNA库来诱发RNAi
互联网
小分子干扰RNA (Small interfering RNA ,siRNA ) 是一种非常有效的工具,能够在包括哺乳动物细胞在内的多个体系中抑制特定基因的表达,从而研究某个基因的功能或者是相关信息。但是这种强有力的方法的难题之一是需要设计,合成siRNA s 和验证其效果,从而找到最有效的siRNA s 。
这个过程需要花费相当的时间和经费。
以化学法合成基因特异的siRNA s 为例,由于设计好的siRNA s 中通常只有大约25% 的siRNA s 能高效抑制基因的表达(抑制效率>80% ),这使得化学合成siRNA 的成本升高。体外转录制备长片断RNA 并不难,但是由于研究表明长片断的双链RNA (>29bps )在哺乳动物细胞中通常会引发抗病毒反应一种非特异基因表达抑制,而目前体外转录往往无法制备小于29bp 的小分子siRNA s 。
一个新的研究发现,用RNase III 降解长片断双链RNA 成为siRNA s 库也能够有效诱导特异的基因沉默,这样可以避免了筛选有效siRNA s 的漫长过程,从而快速,经济的实现特定基因表达的沉默。
在线虫、果蝇和其他一些物种的RNA 干扰(RNA interference ,RNA i) 通路中,长片断的双链RNA 进入细胞后会被一个类似RNase III 的内切核糖核酸酶(Dicer) 降解为一系列的2123bp 长度,3’端有两个碱基突出(带羟基)5’磷酸化的siRNA s 。这些siRNA s 介导同源转录本的降解,从而导致基因表达沉默。( 1-5)
大肠杆菌Escherichia coli RNase III 参与多种细菌RNA s,噬菌体甚至是质粒来源的RNA s 降解(6-9) ,能够将长片断的双链RNA s降解为一系列1215bp 的短双链RNA s,产物末端结构类似Dicer 降解产物(9) ,通过改变RNA seIII 的酶切条件可以提高降解产物的平均长度,达到目前的21bp。
这些21bp的降解产物能够有效介导在哺乳动物细胞和小鼠胚胎中特定基因的RNA i(10,11) 。另外,实验表明,RNase III 的完全降解产物(1215bp)同样可以诱导专一的基因沉默,效果与化学合成或者是酶法得到的siRNA s 相当。
验证RNaseIII 的消化能力
采用RNaseIII 分别消化体外转录制备的人源GAPDH ,La 和c-FOS(200 bp) 双链RNA ,从而验证RNaseIII 的消化能力,结果表明,1个单位的RnaseIII 在37度1小时可以将1mg 的双链RNA 降解为30bp以下,主要是1215bp 的siRNA s 混合物。
用其他基因的双链RNA ,同样证实RNaseIII 能够消化各种双链RNA 序列。另外用Cyclophillin, c-myc, Map Kinase 9, PKC-alpha, Raf-1, Nautilus, 和 h-ras 等基因的双链RNA 作为 RNase III 底物,也可以得到同样的结果。这表明 RNase III 能够消化各种来源的双链RNA 。
验证 RNaseIII 降解产物诱导基因沉默的能力
GAPDH 和La 在Hela细胞中的丰度高,也比较容易检测内源基因的表达水平。将RNaseIII降解GAPDH 和La 得到的siRNA s 库转入Hela 细胞,用免疫荧光法检测其诱导基因沉默的能力。根据所检测的细胞数量对荧光信号进行平衡化和定量。
但是内源的c-FOS 在293细胞中的丰度较低,表达水平下调后就难以检测,为了做同样的验证,实验采用50nM 佛波脂 phorbol ester (PMA) 提前24小时诱导处理使c-FOS 表达水平升高。
用由RNase III 消化c-FOS 双链RNA 得到的siRNA s 混合物转染293细胞,用同样方法检测对基因表达的抑制效果。结果显示用RNase III 消化各种双链RNA 得到的siRNA 库对相应的基因表达抑制效果分别是:GAPDH 表达水平下调78% ,La 表达水平下调86% ,而c-FOS 表达水平则下调75% 。
这个结果表明RNase III 得到的siRNA s 库能够非常有效的抑制基因的表达。

Figure 1. Silencing Gene Targets by RNase III Derived siRNA Cocktails. A 200 bp dsRNA (15 µg) for each gene of interest was digested with 2.5 U RNase III for 1 hour at 37℃. 1A. RNase III efficiently digests dsRNA . One microgram of the dsRNA before and after RNase III digestion was run on a 15% non-denaturing acrylamide gel along with a 21 bp chemically synthesized siRNA to GAPDH, which served as a size marker. The gel was stained with ethidium bromide and photographed under UV light. 1B. RNase III derived siRNA cocktails silence GAPDH, La and c-FOS. GAPDH and La siRNA cocktails were transfected into HeLa cells. The c-fos siRNA mixture was transfected into 293 cells followed by 24 hours of stimulation with 50 nM PMA. All samples were harvested at 48 hours post transfection and immunofluorescence was performed with the appropriate antibodies. Fluorescence signal was quantitated, normalized for cell number and graphed.
12-15bp 的RNase III 降解产物诱导基因沉默的效果
最常用于介导RNA i 的化学合成siRNA 通常是21bp 大小。已经有实验结果证实:用RNase III 在特定条件下降解得到的21bp 大小产物是非常有效的基因表达抑制剂。
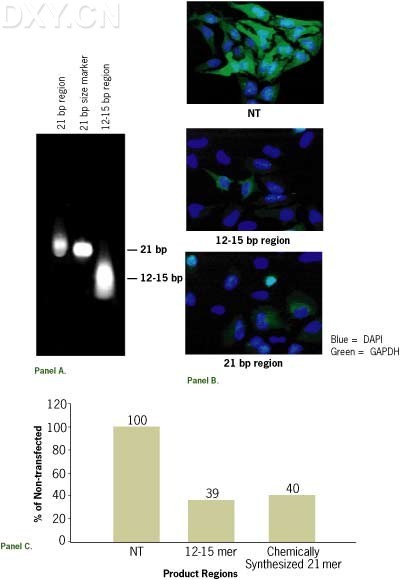
然而RNase III 完全消化产物是1215bp 大小的片断,为了要研究这些12-15bp 大小的片断抑制基因表达的效果,一个200bp 的GAPDH 双链RNA 用RNase III 完全消化得到12-15bp 的产物并用聚丙烯酰胺胶纯化.
按100nM 浓度转染Hela 细胞,另外化学合成的已知能有效抑制GAPDH 表达的siRNA 序列在同样浓度下转染Hela 细胞作为对照,相应的免疫荧光图象表明:12-15bp 的降解产物也能够抑制基因的表达,在相同浓度下两种方法的抑制效果相当。
这个结果表明由RNase III 制备的更短一些的siRNA 混合库能够经过转染哺乳动物细胞抑制靶基因的表达,似乎没有必要通过改变反应条件或者借助纯化手段制备较长的siRNA 从而抑制靶基因的表达。
改变转染的浓度,结果显示siRNA 的转染剂量对基因抑制效果有直接影响,RNase III 制备的siRNA 库需要保持较高的浓度从而达到最大的抑制效果,原因可能是由于:在这个siRNA s 库中既有一些有效的siRNA s 分子,也有很多无效的siRNA s 部分。

Figure 2. 12-15 bp RNase III Digestion Products Elicit Silencing. A 200 bp GAPDH dsRNA (30 µg) was digested with RNase III(30 U) for 1 hour at RT. Digestion products were run on a 15% non-denaturing acrylamide gel and the 12-15 bp products were excised, eluted, and ethanol precipitated. A sample was run on a 15% non-denaturing acrylamide gel for visualization (2A). HeLa cells were transfected with 100 nM of the 12-15 bp RNase III generated GAPDH siRNA s or a 21 bp chemically synthesized GAPDH siRNA . GAPDH protein levels were monitored by immunofluorescence 48 hours after transfection (2B) and the resulting images were quantitated (2C).
基因沉默的专一性
进一步的实验来验证这种方法制备的siRNA 库抑制靶基因表达的专一性:用RNase III 制备的针对GAPDH 的siRNA 库转染Hela 细胞,在降低GAPDH 表达的同时检测其他一系列非特异基因 (La, Ku-70, c-myc, ß-actin, and cdk-2) 的表达水平,结果没有检测到这些非特异基因的表达在转染前后的变化。
结果显示在转染RNase III 制备的siRNA s 库后没有发生非特异基因沉默。 最近一篇关于RNase III 制备的siRNA s 和相关的RNA 结合蛋白的文章同样证实没有非特异基因沉默的发生 (5) 。
在除了哺乳动物细胞以外的其他系统中,可以通过Dicer 酶复合物消化长片断RNA 双链,得到针对同一个靶基因多个位点的siRNA s 群,从而特异的抑制目的基因的表达。这些结果提示存在某种适当的机制来维护抑制反应的高度专一性。
结论
用RNase III 消化dsRNA 制备siRNA 库从而有效抑制目的基因表达,这是一种比较简单粗放、快速、性价比很高的方法。根据实验结果,导入这种方法制备的siRNA 库,并没有表现比用化学合成法制备特定的siRNA s 更高的毒性或者是对基因表达的非特异效应。
采用RNase III 制备siRNA s 混合库可以避免化学合成siRNA 的昂贵费用,siRNA 表达载体的繁琐劳动,以及为找到一个有效的siRNA 序列所必经的漫长的筛选过程,因而成为有别于传统方法的制备siRNA 的另一种好办法,特别是对于只需要运用RNA i 技术作为遗传工具,快速得到RNA 干扰结果,而无需详细研究siRNA 具体信息的研究人员更是如此。

Figure 3. RNase III siRNA Cocktails Show Specificity for Silencing. HeLa cells were transfected with 100 nM RNase III generated siRNA s to GAPDH. Immunofluorescence analysis of GAPDH, La, c-MYC, Cdk-2, Ku-90, and ß-actin was performed 48 hours post transfection and subsequently quantitated.
References
1. Byrom MW, Cheng AM, Ford LP. (2003) Characterizing RNA i induced with siRNA cocktails generated by RNase III. Ambion TechNotes 10(1): 4-6.
2. Yang D, Buchholz F, Huang Z, Goga A, Chen CY, Brodsky FM, Bishop JM. (2002) Short RNA duplexes produced by hydrolysis with Escherichia coli RNase III mediate effective RNA interference in mammalian cells. Proc Natl Acad Sci USA 99(15): 9942-7.
3. Calegari F, Haubensak W, Yang D, Huttner WB, and Bushholz F. (2002) Tissue-specific RNA interference in postimplantation mouse embryos with endoribonuclease-prepared short interfering RNA . Proc Natl Acad Sci USA 99: 14236-40.
4. Trotta R, Vignudilli T, Candini O, Intine RV, Pecorari L, Guerzoni C, santilli G, Byrom MW, Goldoni S, Ford LP, Caligiuri MA, Maraia RJ, Perrotti D, Calabretta B. (2003) Cancer Cell 3: (in press).