MHC抗原的结构及检测
互联网
MHC抗原的结构及检测
MHC编码的抗原为一类同种异体抗原(alloantigen)。其中Ⅰ类和Ⅱ类抗原主要以细胞膜镶嵌蛋白的形式存在,也可脱落成为可溶性的形式。Ⅲ类抗原为补体C2、C4和B因子, 主要分泌到血清等体液中去。
一、MHC抗原的结构
(一)Ⅰ类抗原的结构和分布
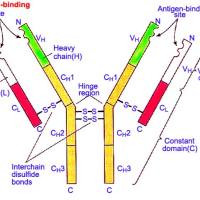
Ⅰ类抗原由非共价键连接的两条多肽链组成,其中重链由MHCⅠ类基因编码,轻链由另一条染色体(人第15对染色体,小鼠第2对染色体)β2m基因编码。 Ⅰ类抗原分布于几乎所有的有核细胞及血小板表面。HLA-A、B抗原在人类淋巴细胞表面浓度最高,每个细胞约有103~105个分子,占淋巴细胞表面蛋白的1%。
1. 重链 又称α链,其裸肽分子量为40kDa。人Ⅰ类抗原α链上有1个N-连接的寡糖, 成熟的α链为糖蛋白,分子量的44kDa;小鼠α链上有2个N-连接的寡糖,分子量略大于人α链,为47kDa。α链为穿膜结构,根据各结构域的功能以及与Ig同源性的比较, α链可分为肽结合区和免疫球蛋白样区组成的胞膜外区,穿膜区以及胞浆区(图6-7)。
(1)肽结合区(peptide-binding region):α链氨基端的两个结构域α1和α2,各含约90氨基酸残基,α1与α2有很高同源性,但不属于免疫球蛋白超家族成员。 人α链有1个N-连接的寡糖,位于α1和α2连接处。小鼠α链则有2个N-连接的寡糖,分别位于α1与α2连接处和α2区的羧基端侧。
α2区内有一个链内的二硫键,两个半胱氨酸间约含63个氨基酸残基。α1区第60~80位氨基酸残基和α2区第 95~120位氨基酸残基的组成和排列顺序变化最大,是Ⅰ类抗原多态性(同种异型)的分子基础。
注: 表示糖,P为磷酸化位点
表示多态性存在部位
美国哈佛大学Strominger 实验室用X射线晶体衍射图搞清了HLA-A2分子的立体结构(图6-8)。α3和β2m结构域靠近细胞膜,位于分子的底部;α1 和α2结构域远离细胞膜位于分子的顶部,所组成的空间结构是与抗原结合部位和被T细胞受体(TCR)识别的部位。
与抗原结合部位的构象呈深槽状,由α1和α2结构域各1条α螺旋和4条β折叠所组成。 两条α螺旋位于抗原结合部位上部形成两个侧面,8条β折叠位于下部形成底面。
所构成的深槽大约长2.5nm,宽1.0nm,深1.1nm,可结合8~20个氨基酸残基,其大小和形状适合于已加工处理的抗原片段。
MHC Ⅰ类抗原分子的多态性主要位于形成两侧面的α螺旋结构上,与Ⅰ类抗原递呈抗原的功能相关,形成深槽内部氨基酸的侧链主要通过盐键、氢键与抗原多肽结合;
位于深槽外部和表面氨基酸是TCR识别的部位, 上述发现是近年来基础免疫学中分子免疫学领域中最杰出的成就之一,为TCR识别MHC与加工处理的抗原复合物的理论研究和某些 自身免疫 性疾病的发病机理提供了重要依据。
(2)免疫球蛋白样区(immunoglobulin-like region):α3约含90个氨基酸残基,氨基酸组成十分保守,与IgC区同源,在二级结构上,α3组成Ig样折叠(Ig fold),即七个β折叠股形成两个平面,由二硫键相连,属免疫球蛋白超家族(IGSF)中C1结构。α3结构域是α重链的非多态部分(nonpolymorphic),通过MHC分子突变分析证实,此区是与CD8分子相互作用的位置。
(3)穿膜区(transmembrane region): α3结构域的羧基端侧有一段较短的连接区(conn-ecting region),穿膜区约由25个疏水性氨基酸残基所组成,可能形成α螺旋穿过双层脂质的细胞膜,并使α链锚在细胞膜上。
(4)胞浆区(cytoplasmic region):含约30个氨基酸残基,并具有数个cAMP依赖的蛋白激酶(蛋白激酶A,PKA)和PP60 Src酪氨酸激酶的磷酸化位点。此外,在羧基端含有一个谷氨酰胺残基,作为转谷氨酰胺酶转肽作用的底物。上述结构可能在MHCⅠ类分子与其它膜蛋白或细胞骨架成分相互作用中起作用,去除MHCⅠ类分子胞浆羧基端可抑制Ⅰ类分子的内化。
2.轻链 含99个氨基酸残基,分子量为12kDa,最早在一些镉中毒患者尿中发现,1968年Berggard 从肾小管病变的蛋白尿中分离出来,电泳位于β2区,称β2微球蛋白(β2-microglobulin, β2m)。A、B、C抗原所含轻链均一致。
β2m氨基酸排列与IgG CH2约有30%序列同源,故β2m有游离免疫球蛋白结构域(free immunoglobulin domain)之称,属免疫球蛋白超家族C1结构。不同种动物之间β2m很少有区别。
β2m本身与HLA-A、B、C抗原特异性无关,也不直接参与同抗原的结合,但对于α重链在细胞膜表面的表达以及执行其正常生理功能是必须的。如人B细胞系Daudi不能合成β2m, 虽然在Dandi细胞Ⅰ类基因能发生转录,并可翻译,但细胞翻译产物是不稳定的。
当Daudi细胞转染了有功能的β2m的基因或与表达β2m的细胞融合,Daudi细胞即可在细胞表面表达Ⅰ类抗原。α重链与β2m的结合可能发生在内织网。有关β2m的免疫学功能近来受到重视。 用抗β2m的单克隆抗体NAMB1可抑制MLR中的反应细胞。
抗原与淋巴细胞接触后18小时内,抗β2m血清可抑制其抗体应答。
此外,β2m抗血清对PHA、ConA、PWM等对淋巴细胞的促有丝分裂作用都有明显的抑制作用。由于β2m分子在细胞表面的数量远远大于Ⅰ类抗原分子的数目,故β2m可能以作为HLAⅠ类抗原一部分和游离分子两种形式存在。在某些病理情况下,血清和尿中β2m可明显升高。
(二)Ⅱ类抗原的结构和分布
1. Ⅱ类分子的结构 MHC Ⅱ类分子是由α、β两条链通过结合紧密的非共价键连接组成的异源双体。α链分子量32~34kDa,有2个N连接寡糖,β链 29~32kDa,有一个N-连接糖基化点(图6-9)。Ⅱ类分子α、β链是由不同基因所碥码。
α、β链各有2个结构域α1、α2及β1、β2。每个结构域约含90氨基酸残基,除α1区外,α2、β1、β2每个区各含一个二硫键。 α1和β1与Ig结构域无相似性,组成肽结合区。α2和β2区与Igγ3和Cμ4相似,属免疫球蛋白超家族C1型结构,非多态性。
所有DR等位基因编码的DR分子α2区结构都相同, 但不同于DPα2或DQα2者。CD4分子与Ⅱ类分子非多态部分相结合。
Ⅱ 类抗原的多态性主要与 DQα、DQβ、DRβ和DPβ链有关。在人类Ⅱ类抗原是HLA-D/DP、DQ、DR抗原,其中D和DP抗原在MLR中为刺激T细胞激活的抗原决定簇 ( major lymphocyte activating determinants,Lads抗原),而DR和DQ则刺激抗体的产生, 这与下述的HLA抗原检测的方法有关。
α2和β2羧基端侧有一个短的连接区。穿膜区约含25个氨基酸残基。胞浆区可能与信号的转导有关。尽管尚未获得α和β链的X-线晶体衍射图,从MHCⅠ、Ⅱ类分子结构和功能的相似性以及最近结果提示MHCⅡ类分子肽结合的多肽折叠形式与MHC Ⅰ类分子十分相似,α1和β1各形成4条β折叠股和一条α螺旋。8条折叠股组成一个底层,支撑2个α螺旋,α、β链2个α螺旋形成肽结合区深槽的侧面。
最近在细胞内发现与MHCⅡ类分子相连的第三条链,称之γ链,约30kDa,属免疫球蛋白超家族成员,非MHC编码,γ链氨基端在胞浆内,而羧基端在细胞膜外。 在γ链到达细胞膜之前它是与MHCⅡ类分子分离的。γ链确切的生理功能还不清楚,可能是:
(1)改变MHCⅡ 类分子α、β链翻译后加工的性质;
(2)参与Ⅱ类分子细胞内的运行;
(3)与Ⅱ类分子将外来抗原提呈给辅助性T细胞的功能有关;
(4)最近又认为γ链可防止机体自身合成的肽与Ⅱ类分子结合,以保证Ⅱ类分子肽结合点为外源性抗原结合之用。
注: 本图为顶面观,白色剪头代表β折叠股,白圈表示α螺旋,黑
色线为二硫键,边缘的虚线方块表示尚未确定的多肽折叠。
2.I区相关抗关
(1)Ia抗原的概念: Klein、Shrefler(1974、1975年) 将小鼠I区基因所编码的抗原称为Ia抗原(I region associated antigen),也有人将I区中的Ir基因的产物称为Ia抗原。目前一般把人类Ⅱ类抗原,尤其DR抗原也称为Ia抗原。
(2)Ia抗原的分布: 小鼠Ia抗原主要在B细胞、巨噬细胞、表皮细胞、精子、活化T细胞、郎罕氏细胞、树突状细胞等。
Ia抗原主要分布在脾脏、淋巴结的淋巴细胞、巨噬细胞、胚胎肝细胞、表皮细胞、活化内皮细胞、骨髓细胞和精子细胞上,某些肿瘤细胞也具有Ia抗原。而红细胞、血小板、脑组织、肾脏和成年肝细胞未见有Ia抗原。
无论是Th还是Tc/Ts被激活后一部分细胞表达Ia抗原。此外巨噬细胞、表皮郎罕氏细胞、树突状细胞也含有较高比例的Ia抗原,有人认为Ia抗原还与巨噬细胞亚群分类有关。
D抗原分布还缺乏系统资料。但从现有资料中看到,除B淋巴细胞外,T细胞、单核细胞、上皮细胞、内皮细胞和精子细胞也可以诱导淋巴母细胞反应,提示上述细胞存在D抗原,而血小板不存在D抗原。