T细胞对同种抗原直接识别的特点和机制
互联网
区别于对一般抗原的应答,由直接识别导致的排斥反应有两个特点:
①速度快,因为无需经历抗原摄取、处理和加工;
②强度大,因为每一个体中,具有同种抗原反应性的T细胞克隆占T细胞库总数的1%一lo%,而针对一般异源性抗原的T细胞克隆仅占总数的1/100 000一l/10 000。
研究表明:
①特异性TCR乃识别pMHC在结合界面所形成的复合结构;
②TCR对pMHC的识别具有简并性(degeneracy),即同一TCR可能识别不同的pMHC;
③CDR3的构型具有包容性(flexibility),可通过构型改变而识别不同pMHC。
上述发现为阐明TCR具有交叉反应性的结构基础以及直接识别的机制,提供了重要依据。
在针对一般抗原的免疫应答中,TCR乃识别外来抗原肽―自身MHC分子所形成的复合结构。在同种移植排斥反应中,受者体内同种反应T细胞所识别的pMHC可能存在两种组合,即外来抗原肽―供者MHC或供者自身肽―供者MHC。
其前提是:抗原肽―供者MHC分子复合物可模拟外来抗原肽―受者MHC分子复合物。换言之,供者APC表面表达多种含供者MHC分子的复合结构(即新表位),它们均可被受者同种反应性T细胞交叉识别。
某些情况下,受者同种反应性T细胞还可直接识别供者MHC分子,而与后者是否结合抗原肽无关(抗原肽的存在仅起稳定MHC分子的作用),此现象称为MHC分子优势结合(MH&dominant binding)。
其机制是:虽然供者MHC分子结构与受者特异性TCR缺乏严格的空间构象互补性,即与受者TCR结合的亲和力低,但由于其高表达于供者APC(及移植物组织细胞)表面,仍可激活受者多克隆T细胞。
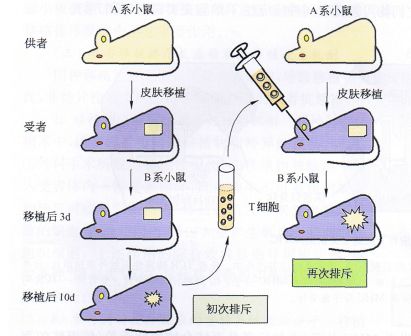
实验研究表明:参与初次移植排斥的同种反应性丁细胞中,许多具有记忆细胞的表型。换言之,数量巨大的参与直接识别、且具有交叉反应性的受者T细胞克隆中,既有接受初次刺激而活化的初始T细胞,也有体内已预存的某些记忆T细胞克隆(后者尤其在急性排斥反应早期发挥作用)。
产生记忆细胞的机制为:正常情况下,受者在移植术前已接触过多种抗原(表位)刺激(如隐性或显性病原微生物感染等),导致识别该外源抗原肽―受者自身MHC分子复合物的T细胞克隆致敏,并形成记忆T细胞;
接受器官移植后,受者体内的记忆T细胞可通过交叉识别机制而识别移植物APC表面的某种供者自身肽/供者MHC分子复合物,并因此被激活。
由于个体生存环境不同,所接触的外源性抗原各异,不同个体体内的记忆T细胞克隆在质(TCR所识别的抗原特异性)和量(接触外源性抗原的种类)上必然存在差异。
不同的供、受者之间,由于遗传背景(主要由MHC多态性决定)和环境因素各异,其体内参与直接识别的记忆T细胞数量、亚群和功能(如与同种抗原肽的亲和力等)也各异,由此导致移植术后不同的转归。