PCR-SSCP原理改进及应用
互联网
随着分子生物学技术的发展,检测基因结构和突变的方法不断涌现。尤其是pCR技术问世以后,各种与pCR相结合的基因检测技术进一步推动了基因研究的发展。如不对称pCR产物的直接测序、核糖核酸酶酶切法(Ribonuclease cleavage,RNAase)、限制性 片段长度多态性分析(Restriction Fragment Length polymorphism,RFLp)等等已成为基因分析的有力工具。
但这些方法操作比较繁琐,局限性较大,或要求实验条件 高,不适合一般临床实验室使用.1989年问世的pCR-SSCp(下文称SSCp)作为检测基因突变的方法,经不断地改进和善,更为简便、快速、灵敏,不但用于检测基因点突 变和短序列的缺失和插入,而且还被用于DNA定量分析,监测pCR诊断实验中的交叉污染情况,以及传染源的调查等.由于SCp的突出的优点,近几年被大量地应用。
一、SSCp的原理及特点
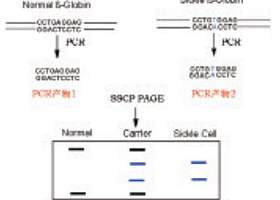
日本Orita等研究发现,单链DNA片段呈复杂的空间折叠构象,这种立体结构主要是由其内部碱基配对等分子内相互作用力来维持的,当有一个碱基发生改变时,会或多或少地影响其空间构象,使构象发生改变,空间构象有差异的单链DNA分子在聚丙烯酰 胺凝胶中受排阻大小不同。
因此,通过非变性聚丙烯酰胺凝胶电泳(pAGE),可以非常 敏锐地将构象上有差异的分子分离开。作者称该方法为单链构象多态性 (Single-Strand Conformation polymorphism,SSCp)分析。在随后的研究中,作者又 将SSCp用于检查pCR扩增产物的基因突变,从而建立了pCR-SSCp技术,进一步提高了 检测突变方法的简便性和灵敏性。其基本过程是:
1、pCR扩增靶DNA;
2、将特异的pCR扩增 产物变性,而后快速复性,使之成为具有一定空间结构的单链DNA分子;
3、将适量的单链DNA进行非变性聚丙烯酰胺凝胶电泳;
4、最后通过放射性自显影、银染或溴化乙锭显色分析结果。
若发现单链DNA带迁移率与正常对照的相比发生改变,就可以判定该链构象发生改变,进而推断该DNA片段中有碱基突变。该方法简便、快速、灵敏,不需要特殊的仪器,适合临床实验的需要。但它也有不足之处。
例如,只能作为一种突变检测方法,要最后确定突变的位置和类型,还需进一步测序;电泳条件要求较严格;另外,由于SSCp是依据点突变引起单链DNA分子立体构象的改变来实现电泳分离的,这样就可能会出现当某些位置的点突变对单链DNA分子立体构象的改变不起作用或作用很小时,再加上其他条件的影响,使聚丙烯酰胺凝胶电泳无法分辨造成漏检。
尽管如此该方法和其他方法相比仍有较高的检测率。首先,它可以发现靶DNA片段中未知位置的碱基突变。
Takao,经实验证明小于300bp的DNA片段中的单碱基突变,90%可被SSCp发现,他认为现在知道的所有单碱基改变绝大多数可用该方法检测出来。另外,SSCp方法可通过聚丙烯酰胺凝胶电泳将不同迁移率的突变单链DNA分离,并且还可以进一步提纯。用这种方法可以最终从DNA序列水平上鉴别突变DNA片段。
二、SSCp的不断改进
SSCp技术自创立以来,经历了自身发展和完善的过程,刚建立时是将同位素掺入pCR扩增物中,通过放射自显影来显示结果。这给该技术的推广造成一定的困难,随着DNA银染方法与pCR-SSCp结合,尤其是直接溴乙锭染色方法的应用,使得该方法大大简化。
最近,SSCp值得注意的改进是将DNA-SSCp分析,改为RNA-SSCp分析,其基本原理是: RNA有着更多精细的二级和三级构象,这些构象对单个碱基的突变很敏感,从而提高 了检出率,其突变检出率可达90%以上。另外,RNA不易结合成双链,因此可以较大量 的进行电泳,有利于用溴化乙锭染色。
但该方法增加了一个反转录过程;还需要一个较 长的引物,内含有启动RNA聚合酶的启动序列,从而相对地增加了该方法的难度。
为了进一步提高SSCp的检出率,可将SSCp分析与其它突变检测方法相结合。其中与杂交双链分析(Heterocluplex analysis,Het)法结合可以大大提高检出率。
Het法是用探针与要检测的单链DNA或RNA进行杂交,含有一对碱基对错配的杂交链可以和完全互 补的杂交链在非变性pAG凝胶上通过电泳被分离开。对同一靶序列分别进行SSCp和Het 分析可以使点突变的检出率接近100%,而且实验简便。
三、pCR-SSCp的实验操作
在小于1Kb长度的情况下,DNA片段长度与丙烯酰胺的浓度选择如下:
DNA片段长度(核苷酸数) (%)
1Kb~700b 3.5
700b~500b 5
500b~200b 8
200b 12
在进行SSCp前首先要通过pCR扩增出特异性好的产物(琼脂糖电泳不能有过强的拖尾)。
1、制备聚丙烯酰胺凝胶(pAG)
按上表制所需的胶液,将梳子插入模子,从梳子一端加入胶,当胶液快到梳齿时,使模子向加胶端倾斜,继续慢慢加胶,边加边放端模子以防止气泡形成,室温放置1h使之凝固。然后拔掉梳子,顶部加入1×TbE封闭,备用。
2、电泳
取10μlpCR产物,加入10μl变性剂(95%甲酰胺,10mmol/LEDTA0.02%溴酚蓝)、30μl石蜡油,煮沸5min,取出立刻放入冰浴中2min以上,然后将水相全部上样,10-15℃下电泳。开始在300V电压下电泳5min,然后在120V电泳8h,取下凝胶,将其浸在含 0.5ug/ml溴化锭的1×TbE缓冲液中染色30--45min,在紫外灯下观察,或进行银染。
3、银染法
将pAG板用去离子水洗二次,浸入10%乙醇和0.5%冰醋酸溶液固定6min。用去离子水洗二次,再浸入0.2%AgNO3溶液中10min。用去离子水洗3--5次,然后浸入1.5%NaOH和0.4%甲醛溶液中显色7min。最后,用0.75%NaCO3终止显色。
四、SSCp的应用
自从Orita等用SSCp进行人类DNA多态性分析以来,该方法大量地用于检测和肿瘤发生有关的基因突变,例如检测星形细胞瘤、脑瘤、小细胞肺癌、胃癌、肠癌等肿瘤中的p53基因的突变、肺癌的ras基因突变等。最近Sugano等用银染色SSCp法,成功地检测了c-Ki-ras2基因第12位的突变,电泳和银染色过程在2.5小时内完成。SSCp还用于检 测引起人类遗传性疾病的研究上,如在囊性纤维化中起作用的CFTR基因、神经纤维瘤 1型基因、家族性结肠息肉基因的研究中。最近,Sharkar等用RNA-SSCp法检测了28名b 型血友病患者的凝血因子IX的基因序列,并与DNA-SSCp和直接基因测序法进行了比 较,发现在全长2.6kbp的凝血因子IX基因组中,有20处碱基点突变,RNA-SSCp可检测 出其中的70%,而DNA-SSCp只能检测出35%,显示了RNA-SSCp比DNA-SSCp有更高的灵敏性。
SSCp法除大量地用于基因突变的检测外,还用于病毒的分型、监测pCR实验中的污染情况、以及病原体传播途径的研究中。Yap用巢式pCR对来自不同国家和地区的HbV标品进行了检测,并对扩增产物进行了SSCp分析,发现每个标本的SSCp图谱完全不同,不仅证明了HbVDNA具有地区性变异,而且排除了交叉性污染的可能性,为保证pCR诊断 结果的可靠性找到了好方法。另外,Yap等还将SSCp用于DNA定量分析上,他们将SSCp 与竞争性pCR法相结合进行乳腺癌细胞突变p53基因的定量分析,并取得了初步成功。 该方法的优点在于,内标和待测DNA只有一个碱基不同,这样使内标和待测DNA有相同 的扩增条件,从而更准确地测出DNA的量。从以上可以看出,SSCp正在分子生物学领域 发挥着巨大的作用。
五、SSCp注意的事项
SSCp是一种快速、简便、灵敏的检测基因突变的方法,为了使SSCp达到最佳效果,应注意下列事项。
1、重复性。影响SSCp重复性的主要因素为电泳的电压和温度。这两个条件保持不变,SSCp图谱可保持良好的重复性。一般SSCp图谱是二条单链DNA带,但有时有的DNA片段可能只呈现一条SSDNA带,或者三条以上,这主要是由于两条单链DNA之间存在相似的立体构象。有时三条以上的SSCp图谱是由于野型DNA片段和突变型DNA片段共同存在的结果。
2、靶DNA序列长度的影响在实验中发现SSCp对短链DNA或RNA的点突变检出率要比长链的高,这可能是由于长链DNA和RNA分子中单个碱基的改变在维持立体构象中起的作用 较小的缘故。而有人认为在DNA链较短的(400bp以下)情况下,DNA的长度不会影响SSCp的效果。他们仔细选择了实验条件,发现354bp的DNA中点突变的检出率仍可达到90%以上。
3、电泳电压和温度的影响:为了使单链DNA保持一定的稳定立体构象,SSCp应在较低温度下进行(一般4℃-15℃之间)。在电泳过程中除环境温度外,电压过高也是引起温度升高的主要原因,因此,在没有冷却装置的电泳槽上进行SSCp时,开始的5min应用较高的电压(250V),以后用100V左右电压进行电泳。这主要是由于开始的高电压可以使 不同立体构象的单链DNA初步分离,而凝胶的温度不会升高。随后的低电压电泳可以使之进一步分离。在实验中应根据具体实验条件确定电泳电压。
4、DNA片段中点突变的位置对SSCp的影响点突变在DNA和RNA中的位置对SSCp检测率的影响,取决于该位置对维持立体构象作用的大小,而不是仅仅取决于点突变在DNA链上的位置(有人认为点突变在DNA链中部要比在近端部容易被SSCp检测出来)。White曾设计了一组样品DNA片段,并将其中的点突变设在DNA链的中部,而且该点又正处在发 夹结构顶端的环上,结果发现SSCp只能检测出9个样品中的2个(检出率为22%),说明 DNA链中任何部位的突变,只要对其单链立体构象没有影响,都有可能被漏检。
5、SSCp的结果断定:由于在SSCp分析中非变性pAG电泳不是根据单链DNA分子和带电量的大小来分离的,而是以单链DNA片段空间构象的立体位阻大小来实现分离的,因此,这种分离不能反映出分子量的大小。有时正常链与突变链的迁移率很接近,很难看出两者之间的差别。因此一般要求电泳长度在16--18cm以上,以检测限为指标来判 定结果。检测限是指突变DNA片段与正常DNA片段可分辨的电泳距离差的最小值。大于检 测限则判定链的迁移率有改变,说明该DNA序列有变化,小于检测限则说明链之间无 变化。例如,一般检测限定为3mm,那么当两带间距离在3mm以上,则说明两链之间有 改变。另外检测限不能定的太低,否则主观因素太大,易造成假阳性结果。另外,SSCp 分析中其它条件,如pCR产物的上样量,pAG的交联度、以及胶的浓度等,都应根据具体实验进行选择确定。总之,SSCp分析法是一种快速、简便、灵敏的突变检测方法, 适合临床实验室的要求。它可以检测各种点突变,短核苷酸序列的缺失或插入。随着SSCp分析不断完善,它将成为基因诊断研究的一个有力工具。