Microarray Analysis of DNA Replication Timing
互联网
DNA replication is a key event in the cell cycle, occurring within a confined period termed S-phase (1 ). Analysis of replication time for individual genes or chromosomal regions has historically relied on either fractionation of S-phase followed by semi-quantitative PCR for synthesis-induced increase in copy number (2 , 3 ) or the counting of FISH signals in S-phase nuclei (4 ). Although, these methods show local variation in replication timing, the laborious nature of the methods have restricted high-resolution analysis to small regions of the chromosomes (3 ) or lower-resolution analysis to single chromosomal segments (2 ). The completion of the human genome sequence and the advent of genome-tiling microarrays have provided an opportunity to study time of replication at a much finer resolution. Here, we detail methods for studying temporal behavior of replication at 25 bp resolution by using nucleotide analog incorporation, density centrifugation, and hybridization. This approach relies on the synchronization of cells to obtain the replication pattern from multiple discrete intervals of S-phase. This strategy has advantages over the existing S:G1 ratio based method of mapping replication timing. In a typical S:G1 method, DNA content is examined at the end and beginning of S-phase and the ratio between these measurements is used to estimate the time of replication. Thus an early replicating segment has a ratio that is closer to 2, while a late replicating segment has a ratio that is closer to 1 (5 ). In such an experiment, segments showing biallelic replication appear to replicate near the middle of S-phase and provide misleading results. In the method described here, computing signal enrichment for multiple time intervals in S-phase allows a finer dissection of the temporal profile of replication and also identifies biallelic or asynchronously replicating regions of the genome more accurately (6 ).
Most genomic methods depend on amplification of the experimental material prior to hybridization; this can change relative amounts of DNA in a complex mixture, and can hence provide unreliable signal values. In the strategy described here, there is no amplification step prior to hybridization, thus freeing the method from any artifacts that could arise due to amplification bias.
We also provide a section in this chapter on the algorithms used to generate continuous TR50 curves along the length of the chromosome. TR50 plots can be used to segregate discrete regions of early, mid, late, and Pan-S replication. Additionally, these curves provide information on genomic regions that undergo acute transition in replication time. These replication domains can be valuable indicators of chromatin structure, as we have observed them to have different levels of gene expression as well as activating and repressing histone marks (6 ). Finally, the local minima of the TR50 curve show areas that replicate earlier than the flanking regions and so are likely to contain origins of replication, as has been shown previously in Saccharomyces cerevisiae , (7 ). Thus, the hundreds of minima in the TR50 profile are likely to be at or near origins of replication.
| 1. | Cell culture: Dulbecco’s Modified Eagle’s Medium (DMEM; CELLGRO) supplemented with 10% iron supplemented donor calf serum (CELLECT) and 1% Penicillin-Streptomycin (GIBCO). |
| 2. | Synchronization: 1 M Thymidine (Sigma) prepared in phosphate-buffered saline (PBS). 1 μg/μl Aphidicolin (Sigma) dissolved in DMSO. |
| 3. | Bromodeoxyuridine (BrdU; Sigma) is dissolved in PBS at 10 mM concentration. Filter sterilized using 0.2-μm filter (CORNING) and stored in aliquots in dark at −20°C. |
| 4. | Propidium iodide FACS: Dilute 1 mg propidium iodide (Sigma) in 10 ml sterile water, 25 μl of 20% NP40, and 10 μl of 10 mg/ml RNase A (Roche Applied Sciences); store wrapped in aluminum foil. |
| 1. | Cell lysis solution: 0.5% SDS, 10 mM Tris-HCl pH 8.0, 300 mM NaCl, 5 mM EDTA, and 200 μg/ml Proteinase K. |
| 2. | Phenol/chloroform/isoamyl-alcohol. |
| 3. | Cholorform/isoamyl-alcohol. |
| 4. | DNase-free ribonuclease (RNase A). |
| 5. | Ethanol, 100% and 70%. |
| 1. | Restriction endonucleases EcoR I and Hind III (NEB). |
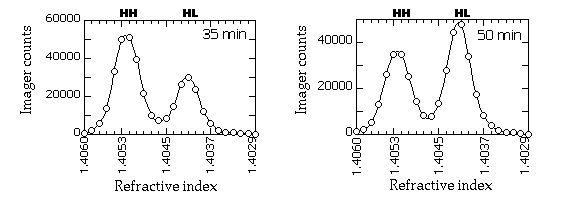
| 2. | CsCl solution: Use 1 g CsCl (Sigma) per ml of TE buffer (10 mM Tris-HCl pH 8.0, 1 mM EDTA pH 8.0). After the CsCl is dissolved in solution, the refractive index should be 1.4052 (see Note 1 ). |
| 3. | OptiSeal tubes (Beckman). |
| 4. | Quick-Seal tubes (Beckman). |
| 1. | 2X SE: 0.8 M NaOH, 20 mM EDTA. |
| 2. | 96-well ELISA plate (BD BioCoat™). |
| 3. | 2 M Ammonium acetate (pH 7.0). |
| 4. | Non-fat dried milk. |
| 5. | Anti-BrdU (Monoclonal antibody to the thymidine-analogue 5-bromo-2′-deoxyuridine Fab fragments with peroxidase (POD) conjugated; Roche Applied Sciences). |
| 6. | TMB substrate (PIERCE). |
| 7. | 2 M H2 SO4 . |
| 1. | DNaseI (Epicenter). |
| 2. | 10X One-Phor-All buffer (Amersham-Pharmacia). |
| 3. | 5X TdT buffer (Roche). |
| 4. | 25 mM CoCl2 (Roche). |
| 5. | 1 mM bio-ddATP (Enzo Life Sciences). |
| 6. | Terminal deoxytransferase (400 U/ml; Roche). |