慢病毒包装系统简介及应用
吉凯基因
一、慢病毒包装简介及其用途
慢病毒(Lentivirus)载体是以 HIV-1 (人类免疫缺陷 I 型病毒)为基础发展起来的基因治疗载体。区别一般的逆转录病毒载体,它对分裂细胞和非分裂细胞均具有感染能力。
慢病毒载体的研究发展得很快,研究的也非常深入。该载体可以将外源基因有效地整合到宿主染色体上,从而达到持久性表达。在感染能力方面可有效地感染神经元细胞、肝细胞、心肌细胞、肿瘤细胞、内皮细胞、干细胞等多种类型的细胞,从而达到良好的基因治疗效果,在美国已经开展了临床研究,效果非常理想,因此具有广阔的应用前景。
目前慢病毒也被广泛地应用于表达 RNAi 的研究中。由于有些类型细胞脂质体转染效果差,转移到细胞内的 siRNA 半衰期短,体外合成 siRNA 对基因表达的抑制作用通常是短暂的,因而使其应用受到较大的限制。采用事先在体外构建能够表达 siRNA 的载体,然后转移到细胞内转录 siRNA 的策略,不但使脂质体有效转染的细胞种类增加,而且对基因表达抑制效果也不逊色于体外合成 siRNA ,在长期稳定表达载体的细胞中,甚至可以发挥长期阻断基因表达的作用。
在所构建的 siRNA 表达载体中,是由 RNA 聚合酶 Ⅲ 启动子来指导 RNA 合成的,这是因为 RNA 聚合酶 Ⅲ 有明确的起始和终止序列,而且合成的 RNA 不会带 poly A 尾。当 RNA 聚合酶 Ⅲ 遇到连续 4 个或 5 个 T 时,它指导的转录就会停止,在转录产物 3' 端形成 1~4 个 U 。 U 6 和 H 1 RNA 启动子是两种 RNA 聚合酶 Ⅲ 依赖的启动子,其特点是启动子自身元素均位于转录区的上游,适合于表达 ~21 ntRNA 和 ~50 ntRNA 茎环结构(stem loop)。
在 siRNA 表达载体中,构成 siRNA 的正义与反义链,可由各自的启动子分别转录,然后两条链互补结合形成 siRNA ;也可由载体直接表达小发卡状 RNA(small hairpin RNA,shRNA),载体包含位于 RNA 聚合酶 Ⅲ 启动子和 4-5 T 转录终止位点之间的茎环结构序列,转录后即可折叠成具有 1~4 个 U 3 ' 突出端的茎环结构,在细胞内进一步加工成 siRNA 。
构建载体前通常要通过合成 siRNA 的方法,寻找高效的 siRNA ,然后从中挑选符合载体要求的序列,将其引入 siRNA 表达载体。
慢病毒载体(Lentiviral vector)较逆转录病毒载体有更广的宿主范围,慢病毒能够有效感染非周期性和有丝分裂后的细胞。慢病毒载体能够产生表达 shRNA 的高滴度的慢病毒,在周期性和非周期性细胞、干细胞、受精卵以及分化的后代细胞中表达 shRNA ,实现在多种类型的细胞和转基因小鼠中特异而稳定的基因表达的功能性沉默,为在原代的人和动物细胞血清中快速而高效地研究基因功能,以及产生特定基因表达降低的动物提供了可能性。
慢病毒表达载体包含了包装、转染、稳定整合所需要的遗传信息。慢病毒包装质粒可提供所有的转录并包装 RNA 到重组的假病毒载体所需要的所有辅助蛋白。
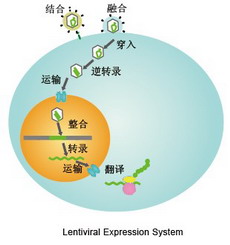
为产生高滴度的病毒颗粒,需要利用表达载体和包装质粒同时共转染细胞,在细胞中进行病毒的包装,包装好的假病毒颗粒分泌到细胞外的培养基中,离心取得上清液后,可以直接用于宿主细胞的感染,目的基因进入到宿主细胞之后,经过反转录,整合到基因组,从而高水平的表达效应分子。
二、这一系统的目的,主要是为了解决以下问题:
- 对于一些较难转染的细胞,如原代细胞、干细胞、不分化的细胞等,能大大提高目的基因转导效率,而且目的基因整合到宿主细胞基因组的几率大大增加,这就为 RNAi,cDNA 克隆以及报告基因的研究提供了一个有利的途径。
- 进行稳转细胞株的筛选;
- 为活体动物模型实验提供高质量的包含目的基因的病毒液;
在细胞相关的实验操作中,对于一些按常规方法难以转染甚至无法转染的细胞,通过病毒介导的实验能够大大提高基因的转导效率,以达到目的基因的高效瞬时表达。
三、慢病毒载体介绍
慢病毒载体(Lentiviral vector,LVs)是在 HIV-1 病毒基础上改造而成的病毒载体系统,它能高效地将目的基因(或 RNAi)导入动物和人的原代细胞或细胞系。
慢病毒载体基因组是正链 RNA ,其基因组进入细胞后,在细胞浆中被其自身携带的反转录酶反转为 DNA,形成 DNA 整合前复合体,进入细胞核后,DNA 整合到细胞基因组中。整合后的 DNA 转录 mRNA ,回到细胞浆中,表达目的蛋白;或产生 RNAi 干扰。
慢病毒载体介导的基因表达或 RNAi 干扰作用持续且稳定,原因是目的基因整合到宿主细胞基因组中,并随细胞基因组的分裂而分裂。
另外,慢病毒载体能有效感染并整合到非分裂细胞中。以上特性使慢病毒载体与其他病毒载体相比,比如不整合的腺病毒载体、整合率低的腺相关病毒载体、只整合分裂细胞的传统逆转录病毒载体,有鲜明的特色。
大量文献研究表明,慢病毒载体介导的目的基因长期表达的组织或细胞包括脑、肝脏、肌肉、视网膜、造血干细胞(HSC)、骨髓间充质干细胞、巨噬细胞等。
慢病毒载体不表达任何 HIV-1 蛋白,免疫原性低,在注射部位无细胞免疫反应,体液免疫反应也较低,不影响病毒载体的第 2 次注射。
四、慢病毒载体的构建原理
慢病毒属于逆转录病毒科,但其基因组结构复杂,除 gag、pol 和 env 这 3 个和单纯逆转录病毒相似的结构基因外,还包括 4 个辅助基因,vif、vpr、nef、vpu 和 2 个调节基因 tat 和 rev 。
HIV 21 是慢病毒中最具特征性的病毒,第一个慢病毒载体系统即以此病毒为基础进行构建的。慢病毒载体的构建原理就是将 HIV 21 基因组中的顺式作用元件 ( 如包装信号、长末端重复序列 ) 和编码反式作用蛋白的序列进行分离。
载体系统包括包装成分和载体成分:包装成分由 HIV 21 基因组去除了包装、逆转录和整合所需的顺式作用序列而构建,能反式提供产生病毒颗粒所需的蛋白;载体成分与包装成分互补,含有包装、逆转录和整合所需的 HIV 21顺式作用序列。同时具有异源启动子控制下的多克隆位点及在此位点插入的目的基因。为降低两种成分同源重组产生有复制能力的病毒 (RCV)的可能性,将包装成分的 5′ LTR 换成巨细胞病毒(CMV)立即早期启动子,3′ LTR 换成 SV 40 polyA 位点等。将包装成分分别构建在两个质粒上,即一个表达 gag 和 pol、另一个表达 env。根据这个原理,Naldini 和 Kafri 等构建了三质粒表达系统。