RNA干扰技术及其应用
丁香园论坛
2004
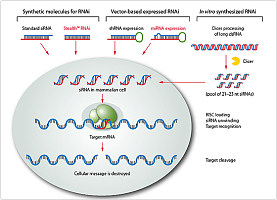
RNA干扰(RNA interference,RNAi)是正常生物体内抑制特定基因表达的一种现象,它是指当细胞中导入与内源性mRNA编码区同源的双链RNA(double stranded RNA,dsRNA)时,该mRNA发生降解而导致基因表达沉默的现象,这种现象发生在转录后水平,又称为转录后基因沉默(post-transcriptional gene silencing,PTGS)。外源dsRNA 进入细胞后产生的小分子干扰RNA(small interfering RNA, siRNA)的反义链和多种核酸酶形成了沉默复合物(RNA-induced silencing complex,RISC),RISC具有结合和切割mRNA的作用而介导RNA干扰的过程。RNAi具有特异性和高效性。这种技术已经成为研究基因功能的重要工具,并将在病毒病、遗传性疾病和肿瘤病的治疗方面发挥重要作用。
近年来,RNA干扰的研究成为热点,预测未来10年间最有可能出重大成果的领域之一,并被《Science》评选为2002年度最重要科技突破的首位。RNAi是细胞本身固有的对抗外源基因及其侵害的一种自我保护现象,是双链RNA(double stranded RNA,dsRNA)分子在mRNA水平关闭相应序列及基因的表达使其沉默的过程。RNAi技术的应用领域已经从基因组学研究逐步扩展到医学领域并作为基因治疗手段。利用RNAi不仅能提供一种经济、快捷、高效的抑制基因表达的技术手段,而且有可能在基因功能测定和基因治疗等方面开辟一条新思路,因此包括动物医学在内的诸多领域的应用也有非常广阔的前景。
1 RNA干扰的发现
1995年,康乃尔大学的Su Guo博士在试图阻断秀丽新小杆线虫(C.elegans)的par-1基因时,发现了一个意想不到的现象。她们本想利用反义RNA技术特异性地阻断上述基因的表达,而同时在对照实验中给线虫注射正义RNA以期观察到基因表达的增强,但得到的结果是二者都同样地切断了par-1基因的表达途径。这是与传统上对反义RNA技术的解释正好相反。该研究小组一直未能给这个意外以合理解释。
1998年,华盛顿卡耐基研究院的Fire和麻省大学医学院的Mello首次在秀丽新小杆线虫中证明上述现象属于转录后水平的基因沉默。他们发现Su Guo博士遇到的正义RNA抑制基因表达的现象,以及过去的反义RNA技术对基因表达的阻断,都是由于体外转录所得RNA中污染了微量双链RNA而引起。当他们将体外转录得到的单链RNA纯化后注射线虫时发现,基因抑制效应变得十分微弱,而经过纯化的双链RNA却正好相反,能够高效特异性阻断相应基因的表达,其抑制基因表达的效率比纯化后的反义RNA至少高2个数量级。该小组将这一现象称为RNA干扰。
在1999年短短的一年间,发现RNA干扰现象广泛存在于从植物、真菌、线虫、昆虫、蛙类、鸟类、大鼠、小鼠、猴一直到人类的几乎所有的真核生物中细胞。2000年,又先后发现小鼠早期胚胎中和大肠杆菌中也存在RNA干扰现象。
2 RNA干扰的作用机制
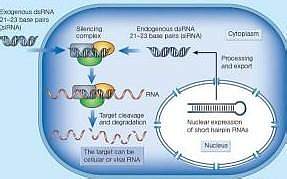
当病毒基因、人工转入基因、转座子等外源性基因随机整合到宿主细胞基因组内,并利用宿主细胞进行转录时,常产生一些dsRNA。宿主细胞对这些dsRNA迅即产生反应,其胞质中的核酸内切酶Dicer将dsRNA切割成多个具有特定长度和结构的小片段RNA(大约21~23 bp),即siRNA。siRNA在细胞内RNA解旋酶的作用下解链成正义链和反义链,继之由反义siRNA再与体内一些酶(包括内切酶、外切酶、解旋酶等)结合形成RNA诱导的沉默复合物(RNA-induced silencing complex,RISC)。RISC与外源性基因表达的mRNA的同源区进行特异性结合,RISC具有核酸酶的功能,在结合部位切割mRNA,切割位点即是与siRNA中反义链互补结合的两端。被切割后的断裂mRNA随即降解,从而诱发宿主细胞针对这些mRNA的降解反应。siRNA不仅能引导RISC切割同源单链mRNA,而且可作为引物与靶RNA结合并在RNA聚合酶(RNA-dependent RNA polymerase,RdRP)作用下合成更多新的dsRNA,新合成的dsRNA再由Dicer切割产生大量的次级siRNA,从而使RNAi的作用进一步放大,最终将靶mRNA完全降解。
RNAi发生于除原核生物以外的所有真核生物细胞内。需要说明的是,由于dsRNA抑制基因表达具有潜在高效性,任何导致正常机体dsRNA形成的情况都会引起不需要的相应基因沉寂。所以正常机体内各种基因有效表达有一套严密防止dsRNA形成的机制。
RNAi干扰现象具有以下几个重要的特征:①RNAi是转录后水平的基因沉默机制;②RNAi具有很高的特异性,只降解与之序列相应的单个内源基因的mRNA;③RNAi抑制基因表达具有很高的效率,表型可以达到缺失突变体表型的程度,而且相对很少量的dsRNA分子(数量远远少于内源mRNA的数量)就能完全抑制相应基因的表达,是以催化放大的方式进行的;④RNAi抑制基因表达的效应可以穿过细胞界限,在不同细胞间长距离传递和维持信号甚至传播至整个有机体以及可遗传等特点;⑤dsRNA不得短于21个碱基,并且长链dsRNA也在细胞内被Dicer酶切割为21 bp左右的siRNA,并由siRNA来介导mRNA切割。而且大于30 bp的dsRNA不能在哺乳动物中诱导特异的RNA干扰,而是细胞非特异性和全面的基因表达受抑和凋亡;⑥ATP依赖性:在去除ATP的样品中RNA干扰现象降低或消失显示RNA干扰是一个ATP依赖的过程。可能是Diecer和RISC的酶切反应必须由ATP提供能量。
3 RNA干扰的应用
RNA干扰现象的发现及其在生物中存在的普遍性,以及RNAi作用机制和生物学功能的初步阐明,为RNAi的应用提供了理论基础。RNAi目前已经在功能基因组学研究、微生物学研究、基因治疗和信号转导等广泛的领域里取得了令人瞩目的进展,使其在医学领域包括动物医学领域在内的应用有着广阔的前景。
3.1 研究基因功能的新工具
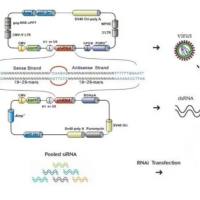
由于RNAi具有高度的序列专一性和有效的干扰活力,可以特异地使特定基因沉默,获得功能丧失或降低突变,因此可以作为功能基因组学的一种强有力的研究工具。已有研究表明RNAi能够在哺乳动物中抑制特定基因的表达,制作多种表型,而且抑制基因表达的时间可以控制在发育的任何阶段,产生类似基因敲除的效应。与传统的基因敲除技术相比,这一技术具有投入少,周期短,操作简单等优势,近来RNAi成功用于构建转基因动物模型的报道日益增多,标志着RNAi将成为研究基因功能不可或缺的工具。
3.2 病毒性疾病的治疗
加州大学洛杉矶分校和加州理工学院的研究人员开发出使用RNAi技术来阻止艾滋病病毒进入人体细胞。这个研究小组设计合成的lenti病毒载体引入siRNA,激发RNAi使其抑制了HIV-1的coreceptor-CCR5进入人体外周T淋巴细胞,而不影响另一种HIV-1主要的coreceptor-CCR4,从而使以lenti病毒载体为媒介引导siRNA进入细胞内产生了免疫应答,由此治疗HIV-1和其他病毒感染性疾病的可行性大大增加。RNAi还可应用于其它病毒感染如脊髓灰质炎病毒等,siRNA已证实介导人类细胞的细胞间抗病毒免疫,用siRNA对Magi细胞进行预处理可使其对病毒的抵抗能力增强。在最近全世界约30个国家和地区散发或流行的严重急性呼吸综合征(severe acute respiratory syndrome,SARS)的防治研究中,RNAi也受到了重视。siRNA在感染的早期阶段能有效地抑制病毒的复制,病毒感染能被针对病毒基因和相关宿主基因的siRNA所阻断,这些结果提示RNAi能胜任许多病毒的基因治疗,RNAi将成为一种有效的抗病毒治疗手段。这对于许多严重的动物传染病的防治具有十分重大的意义。
3.3 遗传性疾病的治疗
美国西北大学的Carthew R W和日本基因研究所的Ishizuka A等人发现RNAi同脆性X染色体综合征(与FMR-1基因异常有关的导致智力低下的染色体病)之间的关系密切,揭示了与RNAi相关机制的缺陷可能导致人类疾病的病理机制。遗传性疾病的RNAi治疗成为当今研究RNAi的又一大热点。
3.4 肿瘤病的治疗
肿瘤是多个基因相互作用的基因网络调控的结果,传统技术诱发的单一癌基因的阻断不可能完全抑制或逆转肿瘤的生长,而RNAi可以利用同一基因家族的多个基因具有一段同源性很高的保守序列这一特性,设计针对这一区段序列的dsRNA分子,只注射一种dsRNA即可以产生多个基因同时剔除的表现,也可以同时注射多种dsRNA而将多个序列不相关的基因同时剔除。Maen等应用RNAi技术成功地阻断了MCF7乳腺癌细胞中一种异常表达的与细胞增殖分化相关的核转录因子基因Sp1的功能。
总之,随着RNAi机制研究的深入和RNA干扰技术的日趋完善,它作为一种便捷实用的基因组研究方法和基因治疗药物,预示着一个崭新的RNA时代即将来临。尽管仍有一些问题亟待研究,如外源转基因如何产生dsRNA?到底有哪些核酸酶参与了dsRNA和mRNA的切割,其详细机制如何?以及最近发现植物的一种钙调素相关蛋白参与PTGS的弱化,提示钙信号可能调节PTGS或RNAi等问题。但细胞中这种古老、多能的细胞水平的监督系统经过千万年的选择和进化的考验,毫无疑问是一件无价之宝。该技术将被广泛地用于农、林、牧、渔等多种领域,具有巨大的科研、经济和社会价值。
近年来,RNA干扰的研究成为热点,预测未来10年间最有可能出重大成果的领域之一,并被《Science》评选为2002年度最重要科技突破的首位。RNAi是细胞本身固有的对抗外源基因及其侵害的一种自我保护现象,是双链RNA(double stranded RNA,dsRNA)分子在mRNA水平关闭相应序列及基因的表达使其沉默的过程。RNAi技术的应用领域已经从基因组学研究逐步扩展到医学领域并作为基因治疗手段。利用RNAi不仅能提供一种经济、快捷、高效的抑制基因表达的技术手段,而且有可能在基因功能测定和基因治疗等方面开辟一条新思路,因此包括动物医学在内的诸多领域的应用也有非常广阔的前景。
1 RNA干扰的发现
1995年,康乃尔大学的Su Guo博士在试图阻断秀丽新小杆线虫(C.elegans)的par-1基因时,发现了一个意想不到的现象。她们本想利用反义RNA技术特异性地阻断上述基因的表达,而同时在对照实验中给线虫注射正义RNA以期观察到基因表达的增强,但得到的结果是二者都同样地切断了par-1基因的表达途径。这是与传统上对反义RNA技术的解释正好相反。该研究小组一直未能给这个意外以合理解释。
1998年,华盛顿卡耐基研究院的Fire和麻省大学医学院的Mello首次在秀丽新小杆线虫中证明上述现象属于转录后水平的基因沉默。他们发现Su Guo博士遇到的正义RNA抑制基因表达的现象,以及过去的反义RNA技术对基因表达的阻断,都是由于体外转录所得RNA中污染了微量双链RNA而引起。当他们将体外转录得到的单链RNA纯化后注射线虫时发现,基因抑制效应变得十分微弱,而经过纯化的双链RNA却正好相反,能够高效特异性阻断相应基因的表达,其抑制基因表达的效率比纯化后的反义RNA至少高2个数量级。该小组将这一现象称为RNA干扰。
在1999年短短的一年间,发现RNA干扰现象广泛存在于从植物、真菌、线虫、昆虫、蛙类、鸟类、大鼠、小鼠、猴一直到人类的几乎所有的真核生物中细胞。2000年,又先后发现小鼠早期胚胎中和大肠杆菌中也存在RNA干扰现象。
2 RNA干扰的作用机制
当病毒基因、人工转入基因、转座子等外源性基因随机整合到宿主细胞基因组内,并利用宿主细胞进行转录时,常产生一些dsRNA。宿主细胞对这些dsRNA迅即产生反应,其胞质中的核酸内切酶Dicer将dsRNA切割成多个具有特定长度和结构的小片段RNA(大约21~23 bp),即siRNA。siRNA在细胞内RNA解旋酶的作用下解链成正义链和反义链,继之由反义siRNA再与体内一些酶(包括内切酶、外切酶、解旋酶等)结合形成RNA诱导的沉默复合物(RNA-induced silencing complex,RISC)。RISC与外源性基因表达的mRNA的同源区进行特异性结合,RISC具有核酸酶的功能,在结合部位切割mRNA,切割位点即是与siRNA中反义链互补结合的两端。被切割后的断裂mRNA随即降解,从而诱发宿主细胞针对这些mRNA的降解反应。siRNA不仅能引导RISC切割同源单链mRNA,而且可作为引物与靶RNA结合并在RNA聚合酶(RNA-dependent RNA polymerase,RdRP)作用下合成更多新的dsRNA,新合成的dsRNA再由Dicer切割产生大量的次级siRNA,从而使RNAi的作用进一步放大,最终将靶mRNA完全降解。
RNAi发生于除原核生物以外的所有真核生物细胞内。需要说明的是,由于dsRNA抑制基因表达具有潜在高效性,任何导致正常机体dsRNA形成的情况都会引起不需要的相应基因沉寂。所以正常机体内各种基因有效表达有一套严密防止dsRNA形成的机制。
RNAi干扰现象具有以下几个重要的特征:①RNAi是转录后水平的基因沉默机制;②RNAi具有很高的特异性,只降解与之序列相应的单个内源基因的mRNA;③RNAi抑制基因表达具有很高的效率,表型可以达到缺失突变体表型的程度,而且相对很少量的dsRNA分子(数量远远少于内源mRNA的数量)就能完全抑制相应基因的表达,是以催化放大的方式进行的;④RNAi抑制基因表达的效应可以穿过细胞界限,在不同细胞间长距离传递和维持信号甚至传播至整个有机体以及可遗传等特点;⑤dsRNA不得短于21个碱基,并且长链dsRNA也在细胞内被Dicer酶切割为21 bp左右的siRNA,并由siRNA来介导mRNA切割。而且大于30 bp的dsRNA不能在哺乳动物中诱导特异的RNA干扰,而是细胞非特异性和全面的基因表达受抑和凋亡;⑥ATP依赖性:在去除ATP的样品中RNA干扰现象降低或消失显示RNA干扰是一个ATP依赖的过程。可能是Diecer和RISC的酶切反应必须由ATP提供能量。
3 RNA干扰的应用
RNA干扰现象的发现及其在生物中存在的普遍性,以及RNAi作用机制和生物学功能的初步阐明,为RNAi的应用提供了理论基础。RNAi目前已经在功能基因组学研究、微生物学研究、基因治疗和信号转导等广泛的领域里取得了令人瞩目的进展,使其在医学领域包括动物医学领域在内的应用有着广阔的前景。
3.1 研究基因功能的新工具
由于RNAi具有高度的序列专一性和有效的干扰活力,可以特异地使特定基因沉默,获得功能丧失或降低突变,因此可以作为功能基因组学的一种强有力的研究工具。已有研究表明RNAi能够在哺乳动物中抑制特定基因的表达,制作多种表型,而且抑制基因表达的时间可以控制在发育的任何阶段,产生类似基因敲除的效应。与传统的基因敲除技术相比,这一技术具有投入少,周期短,操作简单等优势,近来RNAi成功用于构建转基因动物模型的报道日益增多,标志着RNAi将成为研究基因功能不可或缺的工具。
3.2 病毒性疾病的治疗
加州大学洛杉矶分校和加州理工学院的研究人员开发出使用RNAi技术来阻止艾滋病病毒进入人体细胞。这个研究小组设计合成的lenti病毒载体引入siRNA,激发RNAi使其抑制了HIV-1的coreceptor-CCR5进入人体外周T淋巴细胞,而不影响另一种HIV-1主要的coreceptor-CCR4,从而使以lenti病毒载体为媒介引导siRNA进入细胞内产生了免疫应答,由此治疗HIV-1和其他病毒感染性疾病的可行性大大增加。RNAi还可应用于其它病毒感染如脊髓灰质炎病毒等,siRNA已证实介导人类细胞的细胞间抗病毒免疫,用siRNA对Magi细胞进行预处理可使其对病毒的抵抗能力增强。在最近全世界约30个国家和地区散发或流行的严重急性呼吸综合征(severe acute respiratory syndrome,SARS)的防治研究中,RNAi也受到了重视。siRNA在感染的早期阶段能有效地抑制病毒的复制,病毒感染能被针对病毒基因和相关宿主基因的siRNA所阻断,这些结果提示RNAi能胜任许多病毒的基因治疗,RNAi将成为一种有效的抗病毒治疗手段。这对于许多严重的动物传染病的防治具有十分重大的意义。
3.3 遗传性疾病的治疗
美国西北大学的Carthew R W和日本基因研究所的Ishizuka A等人发现RNAi同脆性X染色体综合征(与FMR-1基因异常有关的导致智力低下的染色体病)之间的关系密切,揭示了与RNAi相关机制的缺陷可能导致人类疾病的病理机制。遗传性疾病的RNAi治疗成为当今研究RNAi的又一大热点。
3.4 肿瘤病的治疗
肿瘤是多个基因相互作用的基因网络调控的结果,传统技术诱发的单一癌基因的阻断不可能完全抑制或逆转肿瘤的生长,而RNAi可以利用同一基因家族的多个基因具有一段同源性很高的保守序列这一特性,设计针对这一区段序列的dsRNA分子,只注射一种dsRNA即可以产生多个基因同时剔除的表现,也可以同时注射多种dsRNA而将多个序列不相关的基因同时剔除。Maen等应用RNAi技术成功地阻断了MCF7乳腺癌细胞中一种异常表达的与细胞增殖分化相关的核转录因子基因Sp1的功能。
总之,随着RNAi机制研究的深入和RNA干扰技术的日趋完善,它作为一种便捷实用的基因组研究方法和基因治疗药物,预示着一个崭新的RNA时代即将来临。尽管仍有一些问题亟待研究,如外源转基因如何产生dsRNA?到底有哪些核酸酶参与了dsRNA和mRNA的切割,其详细机制如何?以及最近发现植物的一种钙调素相关蛋白参与PTGS的弱化,提示钙信号可能调节PTGS或RNAi等问题。但细胞中这种古老、多能的细胞水平的监督系统经过千万年的选择和进化的考验,毫无疑问是一件无价之宝。该技术将被广泛地用于农、林、牧、渔等多种领域,具有巨大的科研、经济和社会价值。