RNA干扰技术的原理和应用
互联网
马鹏鹏 薛社普 韩代书
(中国医学科学院基础医学研究所, 中国协和医科大学基础医学院细胞 生物 学系, 北京100005)
RNAi是由双链RNA 所引起的序列特异性基因沉默。RNAi 首先由Fire 等发现于秀丽隐杆线虫(C. elegans) 中, 他们发现将dsRNA 注入线虫体内后可抑制序列同源基因的表达, 并证实这种抑制主要作用于转录之后, 所以又称RNAi 为转录后基因沉默( postt ranscriptional gene silencing ,PTGS) 。
随后人们陆续在果蝇(Drosophila) 、锥虫( Trypanosomes) 、涡虫( Planaria) 、斑马鱼( Zebrafish) 、拟南芥(Arabidopsis thaliana) 、大小鼠和人体内发现了RNAi 现象, 遗传学研究表明RNAi 是真核生物中一种普遍存在且非常保守的机制, 与真核细胞中许多重要生物学过程密切相通过对RNAi 现象的遗传学与生物化学研究,其作用机制已日渐清晰。
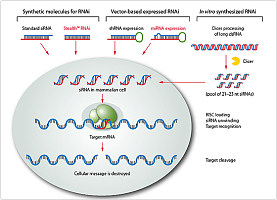
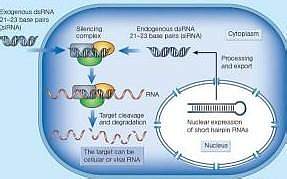
Zamore 等利用果蝇胚胎提取物建立的体外系统, 证实RNAi 是一个依赖ATP 的过程, 在此过程中, dsRNA (外源的或体内产生的) 首先被降解为具5"2单磷酸、长21~23bp 的小分子双链RNA , 这种RNA 分子称为小干扰RNA , siRNA 通过碱基互补配对识别具同源序列的mRNA , 并介导其降解。
研究表明在生物体中siRNA 具相似的结构特征: 为长约21~23bp 的双链RNA , 具5"单磷酸和3"羟基末端, 互补双链的3"端均有一个2~3nt 的单链突出。
在RNAi 过程中一种称为Dicer 的核酸酶负责将dsRNA 转化为siRNA , 它属于RNase Ⅲ家族,具有两个催化结构域、一个解旋酶( helicase) 结构域和一个PAZ ( Piwi/ Argonaute/ Zwille ) 结构域, Dicer 在催化过程中以二聚体的形式出现, 其催化结构域在dsRNA 上反平行排列。
形成四个活性位点, 但只有两侧的两个位点有内切核酸酶活性, 这两个位点在相距约22bp 的距离切断dsRNA , 各种生物体内Dicer 结构略有不同, 致使siRNA 长度存在微小差别。
siRNA 形成之后, 与一系列特异性蛋白结合形成siRNA 诱导干扰复合体( siRNA induced interference complex , RISC) , 此复合物通过碱基互补配对识别靶mRNA 并使其降解, 从而导致特定基因沉默。
在RISC 中, 起靶序列识别作用的是siRNA的反义链 , Zamore 等发现在RNAi 过程中,首先产生的是RISC 无活性前体, 分子量~250kD ,当加入ATP 后可形成100kD 的活性复合体。由无活性前体向活性酶复合物的转换类似蛋白酶原的激要求结合于其上的siRNA 双链的解开。
在ATP 存在时, 依赖于ATP 的解旋酶解开siRNA 的双链并将其正义链与靶mRNA 置换, mRNA 取代正义链与反义链互补, 然后由活化的RISC 在互补区的中间, 距离siRNA 反义链3"末端约12bp 处切断靶mRNA 序列。RNAi 效应具有两个明显的特征, 特异性和高效性。干扰的高效性提示在机制中存在信号放大的步骤。
Fire 等早在1998 年便发现少量的dsRNA就能够导致线虫大量的靶mRNA 降解, 但单凭少量dsRNA 被Dicer 降解为几十个siRNA 并不能解释这种高效性。许多研究显示RNAi 过程中有新的dsRNA 分子的合成, 当siRNA 反义链识别并结合靶mRNA 后。
siRNA 反义链可作为引物, 以靶mRNA 为模板在依赖于RNA 的RNA 聚合酶(RNA2dependent RNA polymerase , RdRP) 催化下合成新的dsRNA , 然后由Dicer 切割产生新的siRNA , 新siRNA 再去识别新一组mRNA , 又产生新的siRNA , 经过若干次合成切割循环, 沉默信号就会不断放大 。正是这种称为靶序列指导的扩增( target2directed amplification) 机制赋予了RNAi 的高效性和持久性。